De RNA eller RNA (ribonukleinsyre) er en type nukleinsyre som er tilstede i eukaryote og prokaryote organismer og i virus. Det er en nukleotidpolymer som inneholder fire typer nitrogenholdige baser i strukturen: adenin, guanin, cytosin og uracil.
RNA er vanligvis funnet som et enkelt bånd (unntatt i noen virus), på en lineær måte eller i en serie av komplekse strukturer. Faktisk har RNA en strukturell dynamikk som ikke blir observert i DNA-dobbeltspiralen. De forskjellige typene RNA har svært varierte funksjoner.
Ribosomale RNA er en del av ribosomer, strukturene som er ansvarlige for proteinsyntese i celler. Messenger-RNA fungerer som mellomledd og transporterer genetisk informasjon til ribosomet, som oversetter meldingen fra en nukleotidsekvens til en aminosyresekvens..
Overførings-RNA er ansvarlig for å aktivere og overføre de forskjellige typene aminosyrer -20 totalt - til ribosomene. Det er et overførings-RNA-molekyl for hver aminosyre som gjenkjenner sekvensen i messenger-RNA.
I tillegg er det andre typer RNA som ikke er direkte involvert i proteinsyntese og deltar i genregulering..
Artikkelindeks
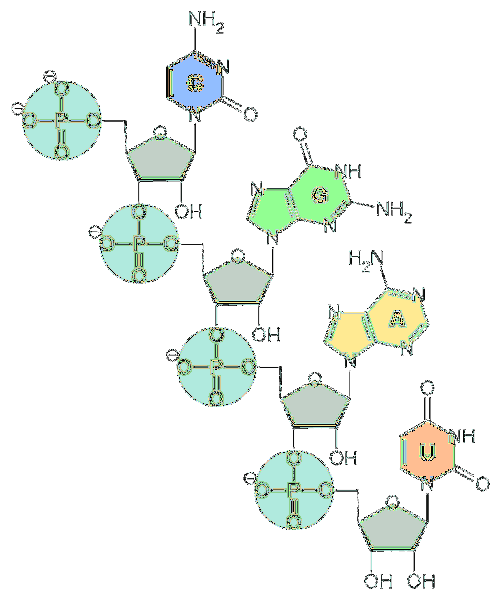
De grunnleggende enhetene til RNA er nukleotider. Hvert nukleotid består av en nitrogenholdig base (adenin, guanin, cytosin og uracil), en pentose og en fosfatgruppe..
Nitrogenbaser er avledet fra to grunnleggende forbindelser: pyrimidiner og puriner..
Basene avledet fra puriner er adenin og guanin, og basene avledet fra pyrimidiner er cytosin og uracil. Selv om dette er de vanligste basene, kan nukleinsyrer også ha andre typer baser som er mindre vanlige..
Når det gjelder pentose, er de enheter av d-ribose. Av denne grunn kalles nukleotidene som utgjør RNA "ribonukleotider".
Nukleotider er bundet sammen av kjemiske bindinger som involverer fosfatgruppen. For å danne dem, er fosfatgruppen ved 5'-enden av et nukleotid festet til hydroksylgruppen (-OH) ved 3'-enden av neste nukleotid, og skaper dermed en fosfodiesterlignende binding..
Langs nukleinsyrekjeden har fosfodiesterbindinger samme orientering. Derfor er det en polaritet av tråden som skiller mellom 3 'og 5' enden.
Etter konvensjon er strukturen til nukleinsyrer representert av 5'-enden til venstre og 3'-enden til høyre.
RNA-produktet av DNA-transkripsjon er et enkeltstrenget bånd som vender mot høyre, i en spiralformet konformasjon ved stabling av baser. Samspillet mellom puriner er mye større enn interaksjonen mellom to pyrimidiner, på grunn av deres størrelse.
I RNA er det ikke mulig å snakke om en tradisjonell og referanse sekundær struktur, som for eksempel den dobbelte helix av DNA. Den tredimensjonale strukturen til hvert RNA-molekyl er unik og kompleks, sammenlignbar med den til proteiner (logisk sett kan vi ikke globalisere strukturen til proteiner).
Det er svake interaksjoner som bidrar til stabilisering av RNA, spesielt base-stabling, der ringene er plassert hverandre. Dette fenomenet bidrar også til stabiliteten til DNA-spiralen..
Hvis RNA-molekylet finner en komplementær sekvens, vil de være i stand til å koble sammen og danne en dobbeltstrenget struktur som vender mot høyre. Den dominerende formen er type A; Når det gjelder Z-skjemaene, har de bare blitt påvist i laboratoriet, mens B-skjemaet ikke er observert.
Generelt er det korte sekvenser (som UUGG) som er lokalisert på enden av RNA og som har det spesielle å danne løkker stabil. Denne sekvensen deltar i brettingen av den tredimensjonale strukturen til RNA.
I tillegg kan hydrogenbindinger dannes på andre steder enn de typiske baseparringene (AU og CG). En av disse interaksjonene forekommer mellom 2'-OH av ribose og andre grupper.
Å belyse de forskjellige strukturene som finnes i RNA har tjent til å demonstrere de mange funksjonene til denne nukleinsyren..
Det er to klasser av RNA: informativ og funksjonell. Den første gruppen inkluderer RNA som deltar i proteinsyntese og fungerer som mellomledd i prosessen; informasjons-RNA er messenger-RNA.
I motsetning til dette, RNA som tilhører andre klasse, de funksjonelle, gir ikke opphav til et nytt proteinmolekyl, og selve RNA er sluttproduktet. Dette er overførings-RNA og ribosomalt RNA..
I pattedyrceller er 80% av RNA ribosomalt RNA, 15% er overførings-RNA, og bare en liten del tilsvarer messenger-RNA. Disse tre typene samarbeider for å oppnå proteinbiosyntese..
Det er også små kjernefysiske RNAer, små cytoplasmatiske RNAer og mikroRNAer, blant andre. Hver av de viktigste typene vil bli beskrevet i detalj nedenfor:
I eukaryoter er DNA begrenset til kjernen, mens proteinsyntese forekommer i cellens cytoplasma, hvor ribosomer finnes. På grunn av denne romlige separasjonen, må det være en formidler som fører meldingen fra kjernen til cytoplasmaet, og at molekylet er budbringer-RNA..
Messenger RNA, forkortet mRNA, er et mellomliggende molekyl som inneholder informasjonen kodet i DNA og som spesifiserer en aminosyresekvens som vil gi opphav til et funksjonelt protein..
Begrepet messenger RNA ble foreslått i 1961 av François Jacob og Jacques Monod for å beskrive den delen av RNA som overførte meldingen fra DNA til ribosomer..
Prosessen med syntese av et mRNA fra DNA-strengen er kjent som transkripsjon, og den skjer forskjellig mellom prokaryoter og eukaryoter..
Genuttrykk styres av flere faktorer og avhenger av behovene til hver celle. Transkripsjon er delt inn i tre trinn: innvielse, forlengelse og avslutning..
DNA-replikasjonsprosessen, som forekommer i hver celledeling, kopierer hele kromosomet. Transkripsjonsprosessen er imidlertid mye mer selektiv, den handler bare om å behandle spesifikke segmenter av DNA-strengen og krever ikke en primer..
På Escherichia coli -den best studerte bakterien innen biovitenskap - transkripsjon begynner med avvikling av DNA-dobbeltspiralen og transkripsjonsløyfen dannes. Enzymet RNA-polymerase er ansvarlig for å syntetisere RNA, og når transkripsjonen fortsetter, går DNA-strengen tilbake til sin opprinnelige form.
Transkripsjon initieres ikke på tilfeldige steder på DNA-molekylet; det er spesialiserte nettsteder for dette fenomenet, kalt promotere. På E coli RNA-polymerase er koblet noen få basepar over målområdet.
Sekvensene der transkripsjonsfaktorer er koblet er ganske konservert mellom forskjellige arter. En av de mest kjente promotorsekvensene er TATA-boksen.
Ved forlengelse tilfører RNA-polymeraseenzymet nye nukleotider til 3'-OH-enden, etter 5 'til 3' -retningen. Hydroksylgruppen fungerer som en nukleofil og angriper alfosfatet av nukleotidet som skal tilsettes. Denne reaksjonen frigjør et pyrofosfat.
Bare en av DNA-strengene brukes til å syntetisere messenger-RNA, som kopieres i 3 'til 5' -retningen (den antiparallelle formen til den nye RNA-strengen). Nukleotidet som skal tilsettes, må være i samsvar med baseparing: U-par med A og G med C.
RNA-polymerase stopper prosessen når den finner regioner som er rike på cytosin og guanin. Etter hvert blir det nye messenger RNA-molekylet skilt fra komplekset.
I prokaryoter kan et messenger-RNA-molekyl kode for mer enn ett protein.
Når et mRNA koder utelukkende for et protein eller polypeptid, kalles det monocistronic mRNA, men hvis det koder for mer enn ett proteinprodukt, er mRNA polycistronic (merk at i denne sammenheng refererer begrepet cistron til genet).
I eukaryote organismer er det store flertallet av mRNAs monocistronic, og transkripsjonsmaskineriet er mye mer komplekst i denne linjen av organismer. De er preget av å ha tre RNA-polymeraser, betegnet I, II og III, hver med spesifikke funksjoner.
Jeg har ansvaret for å syntetisere pre-rRNA, II syntetiserer messenger-RNA og noen spesielle RNA-er. Til slutt er III ansvarlig for overføring av RNA, ribosomal 5S og andre små RNA..
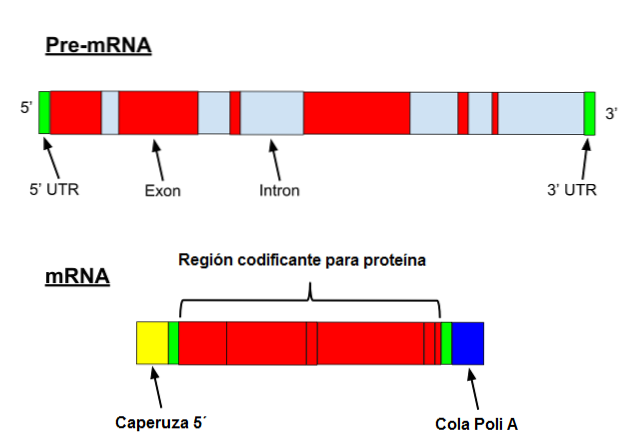
Messenger RNA gjennomgår en rekke spesifikke modifikasjoner i eukaryoter. Den første innebærer å legge til en "cap" til 5'-enden. Kjemisk er hetten en 7-metylguanosinrest festet til enden av en 5 ', 5'-trifosfatbinding..
Funksjonen til denne sonen er å beskytte RNA mot mulig nedbrytning av ribonukleaser (enzymer som bryter ned RNA i mindre komponenter).
I tillegg skjer fjerning av 3'-enden, og 80 til 250 adeninrester tilsettes. Denne strukturen er kjent som polyA "halen" og fungerer som et bindingssted for forskjellige proteiner. Når en prokaryot anskaffer en polyA-hale, har den en tendens til å stimulere nedbrytningen..
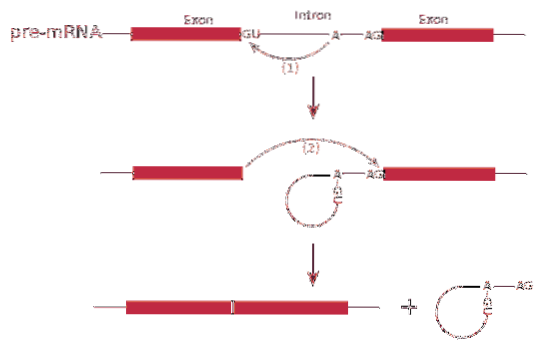
På den annen side er denne senderen transkribert med introner. Introner er DNA-sekvenser som ikke er en del av genet, men som "avbryter" den sekvensen. Introns oversettes ikke og må derfor fjernes fra sendebudet.
De fleste virveldyrgener har introner, med unntak av gener som koder for histoner. Tilsvarende kan antall introner i et gen variere fra noen få til titalls av disse..
Splicing RNA eller skjøtingsprosess innebærer fjerning av introner i messenger RNA.
Noen introner som finnes i nukleære eller mitokondrie gener kan utføre prosessen med skjøting uten hjelp fra enzymer eller ATP. I stedet utføres prosessen ved transesterifiseringsreaksjoner. Denne mekanismen ble oppdaget i ciliated protozoan Tetrahymena thermophila.
Derimot er det en annen gruppe budbringere som ikke er i stand til å megle sine egne skjøting, så de trenger ekstra maskineri. Et ganske stort antall kjernegener tilhører denne gruppen..
Prosessen av skjøting det formidles av et proteinkompleks kalt spliceosome eller spleisingskompleks. Systemet består av spesialiserte RNA-komplekser kalt små kjernefysiske ribonukleoproteiner (RNP)..
Det er fem typer RNP: U1, U2, U4, U5 og U6, som finnes i kjernen og formidler prosessen med skjøting.
De skjøting kan produsere mer enn en type protein - dette er kjent som skjøting alternativt, siden eksonene er ordnet forskjellig, og skaper varianter av messenger-RNA.
Ribosomalt RNA, forkortet rRNA, finnes i ribosomer og deltar i proteinbiosyntese. Derfor er det en viktig komponent i alle celler.
Ribosomalt RNA assosieres med proteinmolekyler (ca. 100) for å gi opphav til ribosomale presubunits. De klassifiseres avhengig av sedimentasjonskoeffisienten, betegnet med bokstaven S for Svedberg-enheter..
Et ribosom består av to deler: den store underenheten og den mindre underenheten. Begge underenhetene skiller seg mellom prokaryoter og eukaryoter når det gjelder sedimentasjonskoeffisienten..
Prokaryoter har en stor 50S-underenhet og en liten 30S-underenhet, mens i eukaryoter er den store underenheten 60S og den lille 40S.
Genene som koder for ribosomale RNA er i nucleolus, et bestemt område av kjernen som ikke er avgrenset av en membran. Ribosomale RNA er transkribert i denne regionen av RNA-polymerase I.
I celler som syntetiserer store mengder proteiner; nucleolus er en fremtredende struktur. Imidlertid, når cellen i spørsmålet ikke krever et stort antall proteinprodukter, er kjernen en nesten umerkelig struktur.
Den 60S store ribosomale underenheten er assosiert med 28S og 5.8S fragmentene. Når det gjelder den lille underenheten (40S), er den assosiert med 18S.
I høyere eukaryoter er pre-rRNA kodet i en 45S-transkripsjonsenhet, som involverer RNA-polymerase I. Dette transkriptet behandles til modne 28S, 18S og 5.8S ribosomale RNA..
Etter hvert som syntese fortsetter, forbinder pre-rRNA med forskjellige proteiner og danner ribonukleoproteinpartikler. Den gjennomgår en serie påfølgende modifikasjoner som inkluderer metylering av 2'-OH-gruppen av ribose og omdannelse av uridinrester til pseudouridin..
Regionen der disse endringene vil forekomme blir kontrollert av mer enn 150 små nukleolære RNA-molekyler, som har evnen til å koble seg til pre-rRNA..
I motsetning til resten av pre-rRNAene blir 5S transkribert av RNA-polymerase III i nukleoplasmaet og ikke inne i nucleolus. Etter å ha blitt syntetisert, blir det ført til nucleolus å montere med 28S og 5.8S, og dermed danner de ribosomale enhetene.
På slutten av monteringsprosessen overføres underenhetene til cytoplasma gjennom kjerneporene..
Det kan skje at et molekyl av messenger RNA gir opphav til flere proteiner samtidig som binder seg til mer enn ett ribosom. Etter hvert som oversettelsesprosessen utvikler seg, blir enden på messenger gratis og kan tas opp av et annet ribosom, og starter en ny syntese..
Av denne grunn er det vanlig å finne ribosomer gruppert (mellom 3 og 10) i et enkelt molekyl av messenger RNA, og denne gruppen kalles polyribosom.
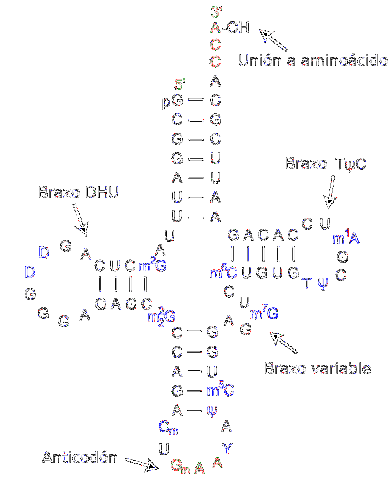
Transfer RNA er ansvarlig for overføring av aminosyrer etter hvert som proteinsyntese prosessen utvikler seg. De består av omtrent 80 nukleotider (sammenlignet med messenger RNA, det er et "lite" molekyl).
Strukturen har bretter og kryss som minner om en trearmet klaver. En adenylisk ring er lokalisert i den ene enden, der hydroksylgruppen av ribose medierer foreningen med aminosyren som skal transporteres.
De forskjellige overførings-RNAene kombineres utelukkende med en av de tjue aminosyrene som utgjør proteiner; med andre ord er det kjøretøyet som bærer byggesteinene til proteiner. Overførings-RNA-komplekset sammen med aminosyren kalles aminoacyl-tRNA.
Videre, i oversettelsesprosessen - som skjer takket være ribosomer - gjenkjenner hvert overførings-RNA et spesifikt kodon i messenger-RNA. Når den gjenkjennes, frigjøres den tilsvarende aminosyren og blir en del av det syntetiserte peptidet.
For å gjenkjenne typen aminosyre som må leveres, har RNA et "antikodon" plassert i midtregionen av molekylet. Dette antikodonet er i stand til å danne hydrogenbindinger med de komplementære basene som er tilstede i messenger DNA..
MicroRNAs eller miRNAs er en type kort, enkeltstrenget RNA, mellom 21 og 23 nukleotider, hvis funksjon er å regulere ekspresjon av gener. Siden det ikke oversettes til protein, kalles det ofte ikke-kodende RNA..
Som andre typer RNA er prosessering av mikroRNA kompleks og involverer et antall proteiner.
MicroRNAs stammer fra lengre forløpere kalt mi-pri-RNA, avledet fra den første transkripsjonen av genet. I cellekjernen modifiseres disse forløperne i mikroprosessorkomplekset, og resultatet er et pre-miRNA..
Pre-miRNAs er 70 nukleotidhårnåler som fortsetter behandlingen i cytoplasmaet av et enzym kalt Dicer, som monterer det RNA-induserte silencing-komplekset (RISC) og til slutt syntetiseres miRNA..
Disse RNAene er i stand til å regulere ekspresjon av gener, siden de er komplementære til spesifikke messenger-RNA. Ved å parre seg med målet deres, er miRNAene i stand til å undertrykke budbringeren, eller til og med nedbryte den. Følgelig kan ikke ribosomet oversette transkripsjonen..
En bestemt type microRNA er små interfererende RNA (siRNA), også kalt dempende RNA. De er korte RNA, mellom 20 og 25 nukleotider, som hindrer ekspresjonen av visse gener.
De er veldig lovende instrumenter for forskning, siden de gjør det mulig å stille et gen av interesse og dermed studere dets mulige funksjon..
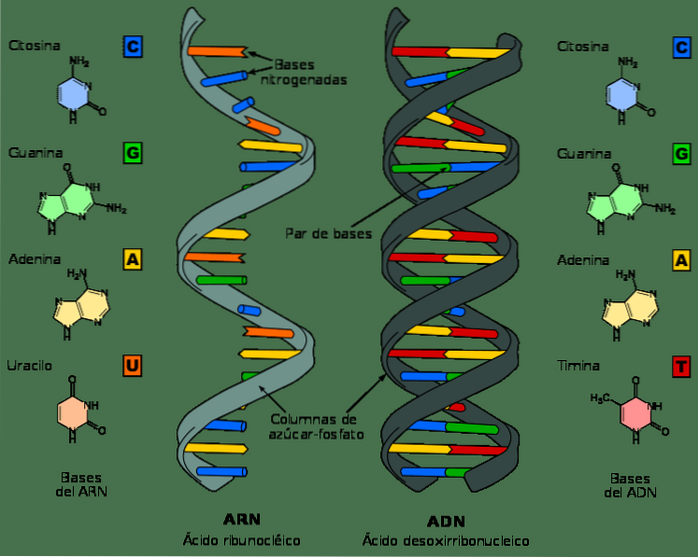
Selv om DNA og RNA er nukleinsyrer og kan se veldig like ut ved første øyekast, varierer de i flere av deres kjemiske og strukturelle egenskaper. DNA er et dobbeltbåndsmolekyl, mens RNA er enkeltbånd.
Som et resultat er RNA et mer allsidig molekyl og kan ta et bredt utvalg av tredimensjonale former. Imidlertid har visse virus dobbeltbånds-RNA i genetisk materiale..
I RNA-nukleotider er sukkermolekylet en ribose, mens det i DNA er en deoksyribose, som bare skiller seg i nærvær av et oksygenatom.
Fosfodiesterbindingen i ryggraden i DNA og RNA er utsatt for en langsom hydrolyseprosess uten tilstedeværelse av enzymer. Under alkaliske forhold hydrolyserer RNA raskt - takket være den ekstra hydroksylgruppen - mens DNA ikke gjør det.
Tilsvarende er de nitrogenholdige basene som utgjør nukleotider i DNA guanin, adenin, tymin og cytosin; derimot, i RNA erstattes tymin med uracil. Uracil kan pares med adenin, på samme måte som tymin i DNA.
RNA er det eneste kjente molekylet som er i stand til å lagre informasjon og katalysere kjemiske reaksjoner samtidig; Av denne grunn foreslår flere forfattere at RNA-molekylet var avgjørende i livets opprinnelse. Overraskende nok er ribosomsubstrater andre RNA-molekyler.
Oppdagelsen av ribozymer førte til biokjemisk omdefinering av "enzym" - siden tidligere ble begrepet eksklusivt brukt for proteiner med katalytisk aktivitet, og bidro til å støtte et scenario der de første livsformene bare brukte RNA som genetisk materiale.



Ingen har kommentert denne artikkelen ennå.