De axoneme Det er en indre cytoskeletal struktur av cilia og flagella basert på mikrotubuli og som gir bevegelse til dem. Dens struktur består av en plasmamembran som omgir et par sentrale mikrotubuli og ni par perifere mikrotubuli..
Aksonemet ligger utenfor cellen og er forankret inne i cellen ved hjelp av basallegemet. Den er 0,2 mikrometer i diameter og lengden kan variere fra 5-10 mikrometer i cilia til flere mm i flagellen av noen arter, selv om disse vanligvis måler 50-150 mikrometer.
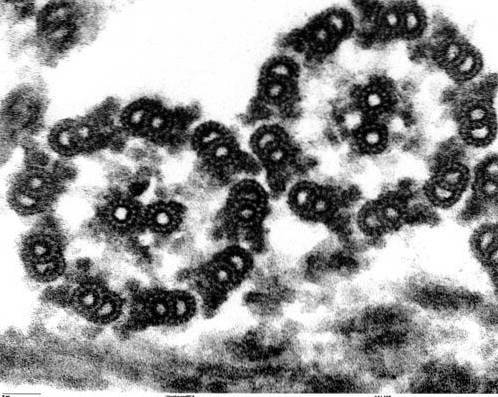
Axonemstrukturen til cilia og flagella er svært konservativ i alle eukaryote organismer, fra mikroalger Chlamydomonas til plagen til den menneskelige sædcellen.
Artikkelindeks
Aksonemene til det store flertallet av cilia og flagella har en konfigurasjon kjent som "9 + 2", det vil si ni par perifere mikrotubuli som omgir et sentralt par.
Mikrotubuli i hvert par er forskjellige i størrelse og sammensetning, bortsett fra det sentrale paret, som presenterer begge mikrotubuli like. Disse rørene er stabile strukturer som er i stand til å motstå brudd..
Mikrotubuli har polaritet, og alle har samme oppsett, med "+" enden mot toppunktet og "-" enden ligger i basen..
Som vi allerede har påpekt, er strukturen til aksonemet av type 9 + 2. Mikrotubuli er lange sylindriske strukturer, som består av protofilamenter. Protofilamenter består i sin tur av proteinunderenheter kalt alfa tubulin og beta tubulin..
Hver protofilament har en alfa tubulin enhet i den ene enden, mens den andre enden har en beta tubulin enhet. Slutten med beta-tubulinterminalen kalles "+" enden, den andre enden vil være "-" enden. Alle protofilamenter i samme mikrotubuli er orientert med samme polaritet.
Mikrotubuli inneholder, i tillegg til tubuliner, proteiner som kalles mikrotubuli-relaterte proteiner (MAP). Av hvert par perifere mikrotubuli består den minste (mikrotubuli A) av 13 protofilamenter.
Mikrotubuli B har bare 10 protofilamenter, men det er større enn mikrotubuli A. Det sentrale paret av mikrotubuli har samme størrelse, og hver av dem består av 13 protofilamenter.
Dette sentrale paret av mikrotubuli er omsluttet av den sentrale kappen, proteinet i naturen, som vil forbinde seg med de perifere A-mikrotubuli ved hjelp av de radiale strålene. På den annen side er mikrotubuli A og B i hvert par sammenføyd av et protein som kalles nexin..
Mikrotubuli En del også et par armer dannet av et protein som kalles dynein. Dette proteinet er ansvarlig for å bruke energien som er tilgjengelig i ATP for å oppnå bevegelse av cilia og flagella.
Eksternt er aksonemet dekket av en ciliær eller flagellær membran som har samme struktur og sammensetning som plasmamembranen i cellen..
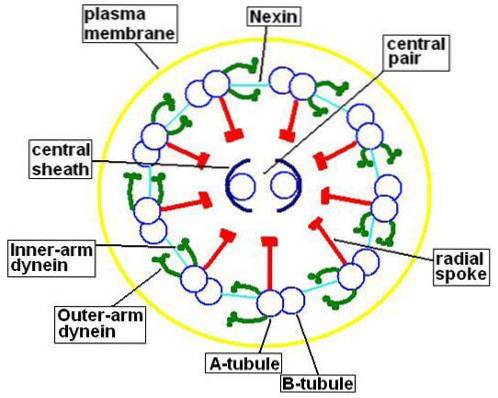
Selv om "9 + 2" -sammensetningen av aksonemet er svært konservert i de fleste eukaryote ciliated og / eller flagellerte celler, er det noen unntak fra denne modellen..
I sædceller fra noen arter går det sentrale paret av mikrotubuli tapt, noe som resulterer i en "9 + 0" -konfigurasjon. Flagellbevegelsen i disse sædcellene ser ikke ut til å variere mye fra den som er observert i aksonemer med normal konfigurasjon, og det antas at disse mikrotubuli ikke spiller en viktig rolle i bevegelsen..
Dette aksonemmønsteret er observert i sædceller fra arter som fisk Lycondontis og av annelider av slekten Myzostomum.
En annen konfigurasjon observert i aksonemer er "9 + 1" -konfigurasjonen. I dette tilfellet er en enkelt sentral mikrotubuli til stede, snarere enn et par. I slike tilfeller er den sentrale mikrotubuli modifisert omfattende, og presenterer flere konsentriske vegger.
Dette aksonememønsteret er blitt observert i hannkjøttene til noen arter av flatorm. Hos disse artene gjentas imidlertid ikke dette axonememønsteret i andre flagellerte eller cilierte celler i organismer..
Studier av flagellabevegelse har vist at flagellafleksjon skjer uten sammentrekning eller forkortelse av mikrotubuli i aksonemet. På grunn av dette har cytologen Peter Satir foreslått en modell for flagellbevegelse basert på forskyvning av mikrotubuli..
I henhold til denne modellen oppnås bevegelse takket være forskyvningen av en mikrotubuli fra hvert par på partneren. Dette mønsteret ligner på glidningen av myosinkjeder på aktin under muskelsammentrekning. Bevegelse skjer i nærvær av ATP.
Dyneinarmene er forankret i mikrotubuli A i hvert par, med endene rettet mot mikrotubuli B. Ved begynnelsen av bevegelse holder dyneinarmene seg til bindingsstedet på mikrotubuli B. Deretter oppstår en endring i konfigurasjonen av dyneinet som driver mikrorør B nedover.
Nexin holder begge mikrorørene nær hverandre. Deretter skilles dyneinarmene fra mikrotubuli B. Deretter vil den bli med på å gjenta prosessen. Denne glidningen skjer vekselvis mellom den ene siden av aksonemet og den andre..
Denne vekslende forskyvningen på den ene siden av aksonemet får cilium, eller flagellum, til å bøyes først til den ene siden og deretter til den motsatte siden. Fordelen med Satirs flagellære bevegelsesmodell er at den ville forklare bevegelsen til vedlegget uavhengig av aksonemkonfigurasjonen til axonem-mikrotubuli.
Det er flere genetiske mutasjoner som kan forårsake unormal utvikling av aksonemet. Disse abnormitetene kan blant annet være mangel på en av dyneinarmene, enten indre eller eksterne, i de sentrale mikrotubuli eller i radiale stråler..
I disse tilfellene utvikler det seg et syndrom kalt Kartageners syndrom, der mennesker som lider av det er infertile fordi sædceller ikke er i stand til å bevege seg.
Disse pasientene utvikler også innvoller i en omvendt stilling i forhold til normal stilling; for eksempel hjertet som ligger på høyre side av kroppen og leveren til venstre. Denne tilstanden er kjent som situs inversus.
De med Kartageners syndrom er også utsatt for luftveis- og bihuleinfeksjoner.
En annen sykdom relatert til unormal utvikling av aksonemet er polycystisk nyresykdom. I dette utvikler det seg flere cyster i nyrene som ender med å ødelegge nyrene. Denne sykdommen skyldes en mutasjon i genene som koder for proteiner som kalles polycystiner.



Ingen har kommentert denne artikkelen ennå.