De replikasjonsgaffel Det er punktet hvor DNA-replikasjon skjer, det kalles også et vekstpunkt. Den er Y-formet, og når replikasjon skjer, beveger hårnålen seg gjennom DNA-molekylet.
DNA-replikasjon er den cellulære prosessen som involverer duplisering av genetisk materiale i cellen. DNA-strukturen er en dobbel helix, og for å replikere innholdet må den åpnes. Hver av strengene vil være en del av den nye DNA-kjeden, siden replikering er en semi-konservativ prosess.
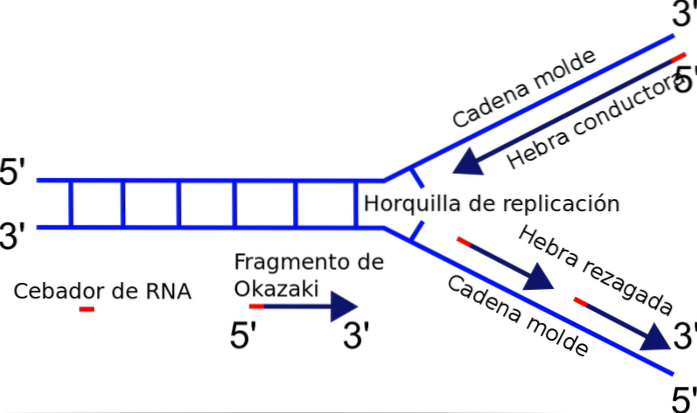
Replikasjonsgaffelen dannes nøyaktig mellom krysset mellom den nylig separerte malen eller malstrengene og dupleks-DNA som ennå ikke er duplisert. Når du starter DNA-replikering, kan en av strengene enkelt dupliseres, mens den andre strengen står overfor et polaritetsproblem..
Enzymet som har ansvaret for polymerisering av kjeden - DNA-polymerase - syntetiserer bare DNA-strengen i 5'-3'-retningen. Dermed er den ene strengen kontinuerlig, og den andre gjennomgår diskontinuerlig replikering og genererer Okazaki-fragmenter..
Artikkelindeks
DNA er molekylet som lagrer den nødvendige genetiske informasjonen til alle levende organismer - med unntak av noen virus.
Denne enorme polymeren består av fire forskjellige nukleotider (A, T, G og C) ligger i kjernen til eukaryoter, i hver av cellene som utgjør vevet til disse vesenene (unntatt i modne røde blodlegemer fra pattedyr, som mangler kjerne).
Hver gang en celle deler seg, må DNA replikere for å skape en dattercelle med genetisk materiale.
Replikering kan være ensrettet eller toveis, avhengig av dannelsen av replikasjonsgaffelen på utgangspunktet..
Logisk sett, når det gjelder replikasjon i en retning, dannes bare en hårnål, mens det i toveis replikasjon dannes to hårnål.
For denne prosessen er det nødvendig med et komplekst enzymatisk maskineri som fungerer raskt og som kan replikere DNA presist. De viktigste enzymene er DNA-polymerase, DNA-primase, DNA-helikase, DNA-ligase og topoisomerase..
DNA-replikering starter ikke noe tilfeldig sted i molekylet. Det er spesifikke regioner i DNA som markerer replikasjonsstart.
I de fleste bakterier har bakteriekromosomet et enkelt AT-rikt startpunkt. Denne sammensetningen er logisk, siden det letter åpningen av regionen (AT-parene er forbundet med to hydrogenbindinger, mens GC-paret med tre).
Når DNA begynner å åpne, dannes en Y-formet struktur: replikasjonsgaffelen..
DNA-polymerase kan ikke starte datterkjedesyntese fra bunnen av. Du trenger et molekyl som har en 3'-ende, slik at polymerasen har hvor du skal begynne å polymerisere.
Denne gratis 3'-enden er levert av et lite nukleotidmolekyl kalt primer. Den første fungerer som en slags krok for polymerasen.
I løpet av replikasjonen har replikasjonsgaffelen muligheten til å bevege seg langs DNA. Gjennomgangen av replikasjonsgaffelen etterlater to enkeltbånds DNA-molekyler som styrer dannelsen av dobbeltbåndsdattermolekylene..
Hårnålen kan gå videre takket være virkningen av helikaseenzymer som avvikler DNA-molekylet. Dette enzymet bryter hydrogenbindinger mellom basepar og tillater hårnålsforskyvning..
Replikeringen er fullført når de to hårnålene er 180 ° C fra opprinnelsen.
I dette tilfellet snakker vi om hvordan replikasjonsprosessen strømmer i bakterier, og det er nødvendig å markere hele torsjonsprosessen til det sirkulære molekylet som replikasjon innebærer. Topoisomeraser spiller en viktig rolle i avviklingen av molekylet.
Har du noen gang lurt på hvordan replikasjon skjer i DNA? Det vil si at en annen dobbel helix må dukke opp fra dobbel helix, men hvordan skjer det? I flere år var dette et åpent spørsmål blant biologer. Flere permutasjoner kan eksistere: to gamle tråder sammen og to nye sammen, eller en ny streng og en gammel streng for å danne den dobbelte helixen..
I 1957 ble dette spørsmålet besvart av forskerne Matthew Meselson og Franklin Stahl. Replikasjonsmodellen foreslått av forfatterne var den semi-konservative.
Meselson og Stahl argumenterte for at resultatet av replikasjon er to DNA-dobbelt-helixmolekyler. Hver av de resulterende molekylene består av en gammel streng (fra det opprinnelige eller opprinnelige molekylet) og en nylig syntetisert ny streng.
DNA-helixen består av to kjeder som går antiparallell: den ene går i 5'-3'-retning og den andre 3'-5 '.
Det mest fremtredende enzymet i replikasjonsprosessen er DNA-polymerase, som er ansvarlig for å katalysere foreningen av de nye nukleotidene som vil bli lagt til kjeden. DNA-polymerase kan bare utvide kjeden i 5'-3'-retning. Dette faktum hindrer samtidig duplisering av kjedene i replikasjonsgaffelen..
Hvorfor? Tilsetningen av nukleotider skjer i den frie enden 3 der en hydroksylgruppe (-OH) er funnet. Således kan bare en av strengene lett amplifiseres ved terminal tilsetning av nukleotidet til 3'-enden. Dette kalles en ledende eller kontinuerlig streng.
Den andre strengen kan ikke være langstrakt, fordi den frie enden er 5 'og ikke 3', og ingen av polymerasene katalyserer tilsetningen av nukleotider til 5'-enden. Problemet løses med syntesen av flere korte fragmenter (130 til 200 nukleotider), hver i normal replikasjonsretning fra 5 'til 3'..
Denne diskontinuerlige syntesen av fragmenter ender med foreningen av hver av delene, en reaksjon katalysert av DNA-ligase. Til ære for oppdageren av denne mekanismen, Reiji Okazaki, kalles de små syntetiserte segmentene Okazaki-fragmenter..



Ingen har kommentert denne artikkelen ennå.