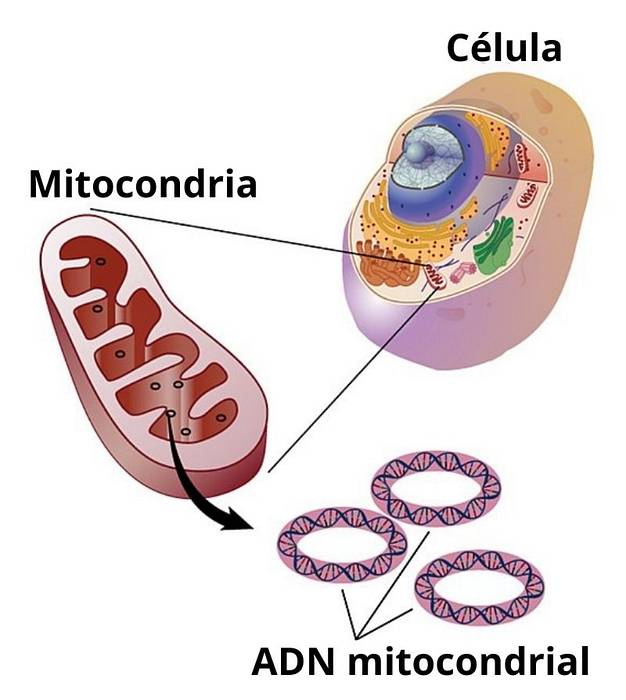
Mitokondrie-DNA er et lite sirkulært DNA-molekyl som finnes inne i disse organellene i eukaryote celler. Dette lille genomet koder for et svært begrenset antall proteiner og aminosyrer i mitokondriene. Det er vanlig å finne navnet "mitokondrie-DNA" forkortet i mange lærebøker og vitenskapelige artikler som "MtDNA"eller på engelsk"mtDNA".
Mitokondrier er essensielle organeller for eukaryote celler, siden de er ansvarlige for å transformere energien fra mat som forbrukes i form av sukker til en form for energi som celler kan bruke (for eksempel ATP).
Alle celler i eukaryote organismer har minst en mitokondrion i seg. Imidlertid er det celler som hjertemuskelceller og skjelettmuskulære celler som kan ha hundrevis av mitokondrier inni seg..
Mitokondrier har sitt eget proteinsynteseapparat uavhengig av celleapparatet, med ribosomer, overførings-RNA og en aminoacyl-RNA-transferase-syntetase fra det indre av organellen; selv om ribosomalt RNA er mindre enn cellen som huser dem.
Et slikt apparat viser stor likhet med apparatet for proteinsyntese av bakterier. Videre, som i prokaryoter, er dette apparatet ekstremt følsomt overfor antibiotika, men veldig forskjellig fra proteinsyntese i eukaryote celler..
Begrepet "mitokondrier" ble introdusert av Benda på slutten av 1100-tallet, og teorien om "endosymbiose" er den mest aksepterte om opprinnelsen. Dette ble publisert i 1967 av Lynn Margulis, i bladet Tidsskrift for teoretisk biologi.
Teorien om "endosymbiose" plasserer mitokondriene for millioner av år siden. Det teoretiseres at en cellulær forfader til eukaryote celler “svelget” og innlemmet en bakterielignende organisme i stoffskiftet, som senere ble det vi nå kjenner som mitokondrier..
Hos pattedyr er generelt hele genomet som inneholder mitokondrie-DNA organisert i et sirkulært kromosom på 15.000 til 16.000 par nukleotider eller, hva er det samme, på 15 til 16 Kb (kilobaser).
Inne i de fleste mitokondrier kan du få flere kopier av mitokondrie-kromosomet. I humane somatiske celler (ikke-kjønnsceller) er det vanlig å finne minst 100 eksemplarer av mitokondrie-kromosomet.
I høyere planter (angiospermer) er mitokondrie-DNA vanligvis mye større, for eksempel, i maisplanten kan det sirkulære kromosomet i mitokondrie-DNA måle opptil 570 Kb.
Mitokondrie-DNA opptar omtrent 1% av det totale DNA-et til de somatiske cellene til de fleste virveldyr. Det er et svært konservert DNA i dyreriket, i motsetning til det som observeres i planter, der det er et stort mangfold.
I noen "gigantiske" eukaryote celler som ovules (kvinnelige kjønnsceller) fra pattedyr eller i celler som inneholder mange mitokondrier, kan mitokondrie-DNA utgjøre opptil 1/3 av totalt cellulært DNA..
Mitokondrie-DNA har noen andre egenskaper enn nukleært DNA: det har en annen tetthet og forhold mellom guanin-cytosin (GC) og adenin-tymin (AT) basepar.
GC basepar tetthet i mitokondrie DNA er 1,68 g / cm3 og innholdet er 21%; mens i kjernefysisk DNA er densiteten 1,68 g / cm3 og innholdet er rundt 40%.
Mitokondrie-DNA har minst 37 gener som er essensielle for normal mitokondriell funksjon. Av de 37 har 13 informasjonen for å produsere enzymene som er involvert i oksidativ fosforylering.
Disse 13 genene koder for 13 polypeptidkomponenter i enzymkompleksene som hører til elektrontransportkjeden og er lokalisert i mitokondriens indre membran..
Til tross for de 13 polypeptidene som mitokondrie-DNA bidrar til elektrontransportkjeden, består den av mer enn 100 forskjellige polypeptider. Imidlertid er disse 13 komponentene essensielle for oksidativ fosforylering og elektrontransportkjeden..
Blant de 13 polypeptidene som er syntetisert fra mitokondrie-DNA, skiller I, II og III-underenhetene til cytokrom C-oksidasekomplekset og VI-underenhet av ATPase-pumper innebygd i organellens indre membran seg ut..
Informasjonen som er nødvendig for syntesen av resten av komponentene som utgjør mitokondriene, er kodet av kjernegener. Disse syntetiseres i cytoplasmaet som resten av cellulære proteiner og importeres deretter til mitokondriene takket være spesifikke signaler..
I oksidativ fosforylering brukes oksygen og sukkeratomer som glukose til syntese eller dannelse av adenosintrifosfat (ATP), som er den kjemiske arten som brukes av alle celler som en energikilde..
De resterende mitokondriegenene har instruksjonene om å syntetisere overførings-RNA (tRNA), ribosomalt RNA og enzymet aminoacyl-RNA-transferase-syntetase (tRNA), nødvendig for syntese av proteiner i mitokondriene..
Inntil relativt nylig ble det antatt at mitokondrie-DNA ble overført utelukkende gjennom mors arv, det vil si direkte nedstigning fra moren..
Imidlertid en artikkel publisert av Shiyu Luo og samarbeidspartnere i bladet Proceedings of the National Academy of Sciences i De forente stater (PNAS) i januar 2019, fant at mitokondrie-DNA i sjeldne tilfeller kan arves fra begge foreldrene, både fra faren og moren.
Før publiseringen av denne artikkelen var det et faktum for forskere at Y-kromosomet og mitokondrie-DNA ble arvet intakt fra henholdsvis far og mor til avkommet..
Den "intakte" arvingen av genene til Y-kromosomet i mitokondrielle gener innebærer at nevnte genetiske materiale ikke gjennomgår endringer ved rekombinasjon, og gjennom årene varierer de bare på grunn av spontane mutasjoner, så variasjonen er ganske lav.
På grunn av dette blir de fleste populasjonsmobiliseringsstudiene gjort på grunnlag av disse genene, for eksempel er det for eksempel enkelt for genetikere å konstruere slektstrær ved bruk av mitokondrie-DNA..
Mye av menneskets historie har blitt rekonstruert gjennom den genetiske historien til mitokondrie-DNA. Mange forretningshus tilbyr til og med å avklare familiebåndet til hver levende person med sine forfedre gjennom teknikker som studerer disse egenskapene..
Den første modellen for mitokondriell DNA-replikering ble foreslått i 1972 av Vinograd og samarbeidspartnere, og denne modellen er fortsatt gyldig, med noen endringer. I det store og hele er modellen basert på en enveis replikering som begynner med to forskjellige replikasjonsopprinnelser..
Forskere klassifiserer mitokondrie-kromosomet i to forskjellige kjeder, den tunge kjeden, H eller OH, fra engelsk "tung"Og lett kjede, L eller OL fra engelsk"Lys”. Disse er identifisert og plassert i de to ikke tildelte åpne leserammene (URF) på mitokondrie-kromosomet.
Replikasjonen av mitokondriegenomet begynner i den tunge kjeden (OH) og fortsetter i en enkelt retning til full lengde på den lette kjeden (OL) er produsert. Deretter er proteiner kalt "mitokondrie enkeltstrengede DNA-bindende proteiner" festet for å beskytte kjeden som fungerer som en "foreldre" eller "mal"..
Enzymer som er ansvarlige for at separasjonen for replikasjon skal forekomme (replikosom) passerer inn i lysbåndet (OL) og det dannes en sløyfestruktur som blokkerer bindingen av mitokondrie enkeltstrengede DNA-bindende proteiner.
I denne sløyfen binder mitokondrie-RNA-polymerase, og syntesen av den nye primeren begynner. Overgangen mot syntesen av den tunge kjeden (OH) skjer 25 nukleotider senere.
Akkurat på tidspunktet for overgangen til den tunge kjeden (OH) erstattes mitokondrie-RNA-polymerase med mitokondrie-replikativ DNA-polymerase i 3'-enden, hvor replikering først begynte.
Til slutt fortsetter syntesen av begge kjedene, både tunge (OH) og lette (OL) kontinuerlig til to komplette sirkulære molekyler av dobbeltstrenget DNA (dobbeltstrenget) dannes..
Det er mange sykdommer relatert til en funksjonsfeil i mitokondrie-DNA. De fleste er forårsaket av mutasjoner som skader sekvensen eller informasjonen i genomet.
En av de best studerte sykdommene som har vært direkte relatert til endringer i mitokondrie DNA genomet er hørselstap på grunn av økende alder.
Denne tilstanden er et produkt av genetiske, miljømessige og livsstilsfaktorer. Når folk eldes, akkumulerer mitokondrie-DNA skadelige mutasjoner, som slettinger, translokasjoner, inversjoner og mer.
Skader på mitokondrie-DNA er hovedsakelig forårsaket av akkumulering av reaktive oksygenarter, disse er biprodukter fra energiproduksjon i mitokondriene.
Mitokondrie-DNA er spesielt utsatt for skade, da det ikke har et reparasjonssystem. Derfor skader endringer forårsaket av reaktive oksygenarter mitokondrie-DNA og fører til at organellen ikke fungerer, og forårsaker celledød..
Cellene i det indre øret har et høyt behov for energi. Dette kravet gjør dem spesielt følsomme for mitokondrie DNA-skader. Disse skadene kan endre funksjonen til det indre øret irreversibelt, og føre til totalt hørselstap..
Mitokondrie-DNA er spesielt følsomt for somatiske mutasjoner, mutasjoner som ikke arves fra foreldrene. Disse typer mutasjoner forekommer i DNA i noen celler gjennom menneskers liv..
Det er bevis som kobler mitokondrie DNA-endringer som følge av somatiske mutasjoner med visse typer kreft, svulster i brystkjertlene, tykktarm, mage, lever og nyre.
Mutasjoner i mitokondrie-DNA har også vært assosiert med blodkreft som leukemi og lymfomer (kreft i immunsystemets celler).
Spesialister knytter somatiske mutasjoner i mitokondrie-DNA med en økning i produksjonen av reaktive oksygenarter, faktorer som øker mitokondrie-DNA-skader og skaper mangel på kontroll i cellevekst.
Det er lite kjent om hvordan disse mutasjonene øker den ukontrollerte celledelingen av celler og hvordan de ender med å utvikle seg som kreftsvulster..
Noen tilfeller av syklisk oppkast, typisk for barndommen, antas å være relatert til mutasjoner i mitokondrie-DNA. Disse mutasjonene forårsaker tilbakevendende episoder med kvalme, oppkast og tretthet eller sløvhet..
Forskere forbinder disse oppkastepisodene med det faktum at mitokondrier med skadet mitokondrie-DNA kan påvirke visse celler i det autonome nervesystemet og påvirke funksjoner som hjertefrekvens, blodtrykk og fordøyelse..
Til tross for disse assosiasjonene er det ennå ikke klart hvordan endringer i mitokondrie-DNA forårsaker tilbakevendende episoder av syklisk oppkastssyndrom..


Ingen har kommentert denne artikkelen ennå.