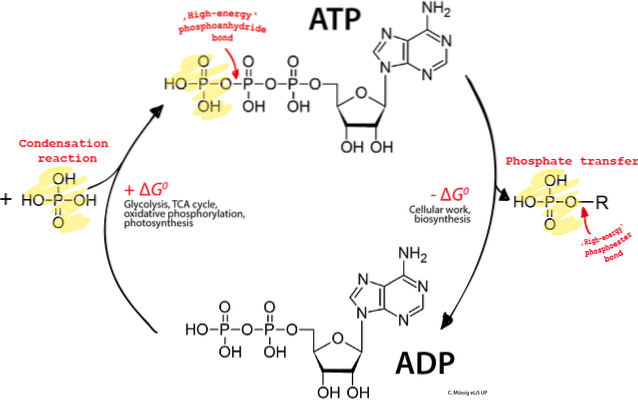
De ATP (adenosintrifosfat) er et organisk molekyl med høyenergibindinger som består av en adeninring, en ribose og tre fosfatgrupper. Den har en grunnleggende rolle i stoffskiftet, siden den transporterer energien som er nødvendig for å holde en rekke cellulære prosesser som fungerer effektivt..
Det er allment kjent under begrepet "energivaluta", siden det dannes og brukes lett, slik at "raskt" kan betales for kjemiske reaksjoner som krever energi.

Selv om molekylet med det blotte øye er lite og enkelt, lagrer det en betydelig mengde energi i sine bindinger. Fosfatgrupper har negative ladninger, som er i konstant frastøting, noe som gjør det til en labil og lett ødelagt binding..
Hydrolysen av ATP er nedbrytningen av molekylet ved tilstedeværelse av vann. Ved denne prosessen frigjøres den inneholdte energien.
Det er to hovedkilder til ATP: fosforylering av substratnivå og oksidativ fosforylering, sistnevnte er den viktigste og mest brukte av cellen..
Oksidativ fosforylering kobler oksidasjon av FADHto og NADH + H+ i mitokondrier og substratnivå oppstår fosforylering utenfor elektrontransportkjeden, i veier som glykolyse og trikarboksylsyresyklusen.
Dette molekylet er ansvarlig for å gi den energien som er nødvendig for de fleste av prosessene som skjer inne i cellen for å finne sted, fra proteinsyntese til bevegelse. I tillegg tillater det trafikk av molekyler gjennom membraner og virker på cellesignalering..
Artikkelindeks
ATP, som navnet antyder, er et nukleotid med tre fosfater. Den spesielle strukturen, spesielt de to pyrofosfatbindinger, gjør den til en energirik forbindelse. Den består av følgende elementer:
- En nitrogenholdig base, adenin. Nitrogenbaser er sykliske forbindelser som inneholder ett eller flere nitrogen i strukturen. Vi finner dem også som komponenter i nukleinsyrer, DNA og RNA.
- Ribose ligger i sentrum av molekylet. Det er et sukker av pentose-typen, siden det har fem karbonatomer. Dens kjemiske formel er C5H10ELLER5. Karbon 1 av ribose er festet til adeninringen.
- Tre fosfatradikaler. De to siste er "høyenergibindinger" og er representert i de grafiske strukturene av vippesymbolet: ~. Fosfatgruppen er en av de viktigste i biologiske systemer. De tre gruppene kalles alfa, beta og gamma, fra nærmeste lengst.
Denne lenken er veldig labil, så den deler seg raskt, enkelt og spontant når organismenes fysiologiske forhold tilsier det. Dette skjer fordi de negative ladningene til de tre fosfatgruppene hele tiden prøver å bevege seg bort fra hverandre..
ATP spiller en uunnværlig rolle i energimetabolismen til praktisk talt alle levende organismer. Av denne grunn blir det ofte referert til som en energivaluta, ettersom den kontinuerlig kan brukes og etterfylles på få minutter..
Direkte eller indirekte gir ATP energi til hundrevis av prosesser, i tillegg til å fungere som en fosfatgiver.
Generelt fungerer ATP som et signalmolekyl i prosessene som forekommer inne i cellen, det er nødvendig å syntetisere komponentene i DNA og RNA, og for syntese av andre biomolekyler deltar det i handel gjennom blant annet.
Bruken av ATP kan deles inn i hovedkategorier: transport av molekyler gjennom biologiske membraner, syntesen av forskjellige forbindelser og til slutt mekanisk arbeid.
Funksjonene til ATP er veldig brede. Videre er det involvert i så mange reaksjoner at det ville være umulig å nevne dem alle. Derfor vil vi diskutere tre spesifikke eksempler for å eksemplifisere hver av de tre nevnte bruksområdene..
Cellen er et svært dynamisk miljø som krever at spesifikke konsentrasjoner opprettholdes. De fleste molekyler kommer ikke tilfeldig eller tilfeldig inn i cellen. For at et molekyl eller stoff skal komme inn, må det gjøre det av sin spesifikke transportør.
Transportører er membranomspennende proteiner som fungerer som cellulære "portvoktere", som kontrollerer materialstrømmen. Derfor er membranen semi-permeabel: den tillater at visse forbindelser kommer inn og andre ikke..
En av de mest kjente transportene er natrium-kaliumpumpen. Denne mekanismen er klassifisert som aktiv transport, siden ioners bevegelse skjer mot deres konsentrasjoner, og den eneste måten å utføre denne bevegelsen på er å innføre energi i systemet, i form av ATP..
Det anslås at en tredjedel av ATP dannet i cellen brukes til å holde pumpen aktiv. Natriumioner pumpes konstant til utsiden av cellen, mens kaliumioner pumpes i motsatt retning..
Logisk er bruken av ATP ikke begrenset til transport av natrium og kalium. Det er andre ioner, for eksempel kalsium, magnesium, som trenger denne energivalutaen for å komme inn.
Proteinmolekyler består av aminosyrer, bundet sammen av peptidbindinger. For å danne dem kreves det å bryte fire høyenergibindinger. Med andre ord må et betydelig antall ATP-molekyler hydrolyseres for dannelse av et protein med gjennomsnittlig lengde..
Proteinsyntese forekommer i strukturer som kalles ribosomer. Disse er i stand til å tolke koden som messenger RNA har og oversette den til en aminosyresekvens, en ATP-avhengig prosess..
I de mest aktive cellene kan proteinsyntese lede opptil 75% av ATP syntetisert i dette viktige arbeidet.
På den annen side syntetiserer cellen ikke bare proteiner, den trenger også lipider, kolesterol og andre essensielle stoffer, og for å gjøre det krever den energien i ATP-bindingene..
Mekanisk arbeid er en av de viktigste funksjonene til ATP. For eksempel for at kroppen vår skal kunne trekke muskelfibre, er tilgjengeligheten av store mengder energi nødvendig..
I muskler kan kjemisk energi transformeres til mekanisk energi takket være omorganiseringen av proteinene med kapasitet til å trekke seg sammen. Lengden på disse strukturene er modifisert, forkortet, noe som skaper en spenning som oversettes til bevegelsesgenerering.
I andre organismer forekommer bevegelse av celler også takket være ATP. For eksempel forekommer bevegelse av cilia og flagella som tillater bevegelse av visse encellede organismer ved bruk av ATP.
En annen spesiell bevegelse er den amøbeiske som involverer et fremspring av en pseudopod ved celleendene. Flere celletyper bruker denne bevegelsesmekanismen, inkludert leukocytter og fibroblaster..
Når det gjelder kimceller, er bevegelse viktig for en effektiv utvikling av embryoet. Embryonale celler beveger seg viktige avstander fra opprinnelsesstedet til regionen der de må stamme fra spesifikke strukturer.
Hydrolysen av ATP er en reaksjon som involverer nedbrytningen av molekylet ved tilstedeværelse av vann. Reaksjonen er representert som følger:
ATP + Vann ⇋ ADP + PJeg + Energi. Hvor, begrepet PJeg refererer til den uorganiske fosfatgruppen og ADP er adenosindifosfat. Merk at reaksjonen er reversibel.
Hydrolysen av ATP er et fenomen som involverer frigjøring av en enorm mengde energi. Brudd på noen av pyrofosfatbindinger resulterer i frigjøring av 7 kcal per mol - spesielt 7,3 fra ATP til ADP og 8,2 for produksjon av adenosinmonofosfat (AMP) fra ATP. Dette tilsvarer 12 000 kalorier per mol ATP..
Fordi hydrolyseproduktene er mye mer stabile enn den opprinnelige forbindelsen, det vil si ATP.
Det bør nevnes at bare hydrolysen som oppstår på pyrofosfatbindinger for å gi opphav til dannelsen av ADP eller AMP fører til generering av energi i betydelige mengder.
Hydrolysen av de andre bindingene i molekylet gir ikke så mye energi, med unntak av hydrolyse av uorganisk pyrofosfat, som har en stor mengde energi.
Frigjøring av energi fra disse reaksjonene brukes til å utføre metabolske reaksjoner inne i cellen, siden mange av disse prosessene trenger energi for å fungere, både i de første trinnene av nedbrytningsveiene og i biosyntese av forbindelser..
For eksempel, i glukosemetabolisme, involverer de innledende trinnene fosforylering av molekylet. I de følgende trinnene genereres ny ATP for å oppnå et positivt nettofortjeneste.
Fra energisynspunktet er det andre molekyler hvis frigjøringsenergi er større enn ATP, inkludert 1,3 bisfosfoglyserat, karbamylfosfat, kreatininfosfat og fosfoenolpyruvat..
ATP kan oppnås på to måter: oksidativ fosforylering og fosforylering på substratnivå. Førstnevnte krever oksygen mens sistnevnte ikke. Omtrent 95% av den dannede ATP forekommer i mitokondriene.
Oksidativ fosforylering involverer en to-fase næringsoksydasjonsprosess: å oppnå reduserte koenzymer NADH og FADHto stammer fra vitaminer.
Reduksjonen av disse molekylene krever bruk av hydrogen fra næringsstoffer. I fett er produksjonen av koenzymer bemerkelsesverdig, takket være den enorme mengden hydrogener de har i strukturen, sammenlignet med peptider eller karbohydrater.
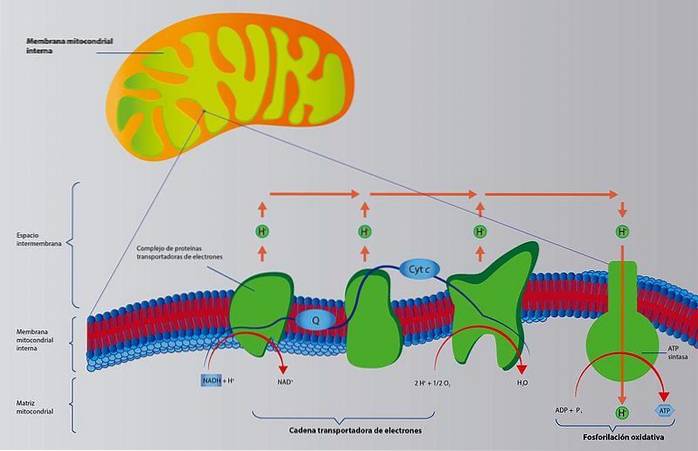
Selv om det er flere ruter for koenzymproduksjon, er den viktigste ruten Krebs-syklusen. Deretter konsentreres de reduserte koenzymer i luftveiskjedene i mitokondriene, som overfører elektronene til oksygen..
Elektrontransportkjeden består av en serie membrankoblede proteiner som pumper protoner (H +) utover (se bilde). Disse protonene kommer inn og krysser membranen igjen ved hjelp av et annet protein, ATP-syntase, som er ansvarlig for syntesen av ATP.
Med andre ord har vi at reduksjonen av koenzymer, mer ADP og oksygen genererer vann og ATP.
Fosforylering på substratnivå er ikke like viktig som mekanismen beskrevet ovenfor, og siden den ikke krever oksygenmolekyler, er den ofte assosiert med gjæring. Selv om den er veldig rask, trekker den ut lite energi, hvis vi sammenligner den med oksidasjonsprosessen, vil den være omtrent femten ganger mindre.
I kroppen vår skjer gjæringsprosesser på muskelnivå. Dette vevet kan fungere uten oksygen, så det er mulig at et glukosemolekyl blir nedbrutt til melkesyre (for eksempel når vi gjør en del uttømmende sportsaktivitet).
I gjæringer har sluttproduktet fortsatt energipotensial som kan ekstraheres. I tilfelle gjæring i muskler er karbonene i melkesyre på samme reduksjonsnivå som de i det opprinnelige molekylet: glukose.
Dermed skjer energiproduksjon gjennom dannelsen av molekyler som har høyenergibindinger, inkludert 1,3-bisfosfoglyrat og fosfoenolpyruvat..
I glykolyse, for eksempel, er hydrolysen av disse forbindelsene knyttet til produksjonen av ATP-molekyler, derav begrepet "på substratnivå".
ATP lagres aldri. Det er i en kontinuerlig syklus av bruk og syntese. Dette skaper en balanse mellom det dannede ATP og dets hydrolyserte produkt, ADP..
ATP er ikke det eneste molekylet som består av bisfosfatnukleosider som eksisterer i cellulær metabolisme. Det er en rekke molekyler med strukturer som ligner på ATP som har sammenlignbar energeatferd, selv om de ikke er like populære som ATP.
Det mest fremtredende eksemplet er GTP, guanosintrifosfat, som brukes i den velkjente Krebs-syklusen og i den glukoneogene banen. Andre mindre brukte er CTP, TTP og UTP.


Ingen har kommentert denne artikkelen ennå.