De natriumkaliumpumpe Det er en proteinstruktur som inngår i et bredere sett med molekyler som finnes i mange cellemembraner, og som er ansvarlig for den aktive transporten av ioner eller andre små molekyler mot konsentrasjonsgradientene. De bruker energien som frigjøres ved hydrolyse av ATP, og det er derfor de generelt kalles ATPaser.
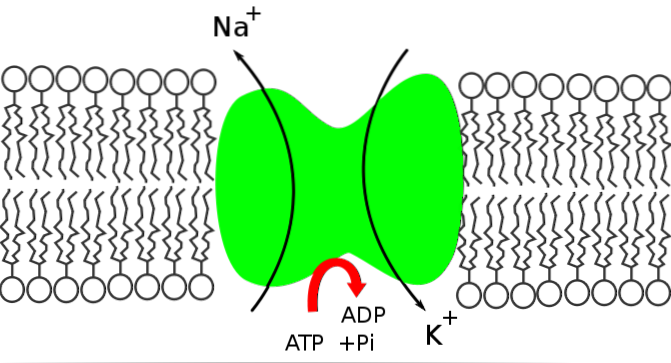
Natriumkaliumpumpen er en Na + / K + ATPase fordi den frigjør energien i ATP-molekylet for å flytte natrium fra innsiden til utsiden av cellen, mens den introduserer kalium.
Inne i cellen er natrium mindre konsentrert (12 mEq / L) enn utenfor (142 mEq / L), mens kalium er mer konsentrert utenfor (4 mEq / L) enn innsiden (140 mEq / L).
ATPase-pumper er klassifisert i tre store grupper:
Natriumkaliumpumpen (Na + / K + ATPase) tilhører gruppen av ionepumper i P-klasse og ble oppdaget i 1957 av Jens Skou, en dansk forsker, da han studerte virkningsmekanismen til anestetika på nervene til en krabbe. (Carcinus maenas); arbeid som han ble tildelt Nobelprisen i kjemi i 1997.
Artikkelindeks
Natriumkaliumpumpen er et enzym som fra synspunktet av den kvaternære strukturen er dannet av 2 proteinunderenheter av alfa (α) -typen og to av beta (β) -typen.
Det er derfor en tetramer av α2β2-typen, hvis underenheter er integrerte membranproteiner, det vil si at de krysser lipid-dobbeltlaget og har både intra- og ekstra-cytosoliske domener..
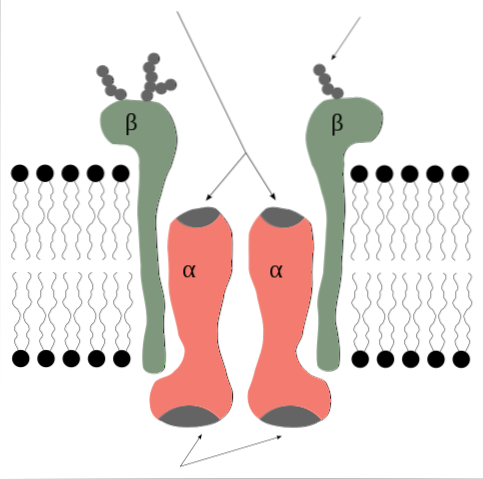
Α-underenhetene er de som inneholder bindingsstedene for ATP og for Na + og K + -ionene og representerer den katalytiske komponenten i enzymet og den som utøver selve pumpens funksjon..
Α-underenhetene er store polypeptider, med en molekylvekt på 120 kDa, 10 transmembrane segmenter og med deres N- og C-terminale ender plassert på den cytosoliske siden..
På den intracellulære siden har de bindingssteder for ATP og Na +, samt en aspartatrest i posisjon 376 som representerer stedet som gjennomgår fosforyleringsprosessen under pumpeaktivering..
Bindingsstedet for K + ser ut til å være på den ekstracellulære siden.
Β-underenhetene ser ikke ut til å ha en direkte deltakelse i pumpefunksjonen, men i deres fravær oppstår ikke denne funksjonen..
P-underenhetene har en molekylvekt på ca. 55 kDa hver og er glykoproteiner med et enkelt transmembrandomene hvis karbohydratrester er satt inn i det ekstracellulære området..
De ser ut til å være nødvendige i det endoplasmatiske retikulumet, hvor de vil bidra til riktig folding av α-underenhetene, og deretter, på membrannivå, for å stabilisere komplekset..
Begge typer underenheter er heterogene, og α1, α2 og α3 isoformer er beskrevet for den ene, og β1, β2 og β3 for den andre. Α1 finnes i membranene i de fleste celler, mens α2 er tilstede i muskel, hjerte, fettvev og hjerne og α3 i hjerte og hjerne.
Β1-isoformen er den med den mest diffuse distribusjonen, selv om den er fraværende i noen vev som vestibulære celler i det indre øret og de raskt reagerende glykolytiske muskelcellene. Sistnevnte inneholder bare β2.
De forskjellige strukturene til underenhetene som utgjør Na + / K + -pumpen i de forskjellige vevene, kan skyldes funksjonelle spesialiseringer som ennå ikke er belyst..
For ethvert øyeblikk utgjør plasmamembranen en separasjonsgrense mellom rommet som tilsvarer det indre av en celle og det som representerer det ekstracellulære væsken det er nedsenket i..
Begge rom har en sammensetning som kan være kvalitativt forskjellig, siden det er stoffer inne i celler som ikke finnes utenfor dem, og den ekstracellulære væsken inneholder stoffer som ikke er tilstede intracellularly..
Stoffer som er tilstede i begge avdelinger kan finnes i forskjellige konsentrasjoner, og disse forskjellene kan ha fysiologisk betydning. Slik er tilfellet med mange ioner.
Na + / K + -pumpen spiller en grunnleggende rolle for å opprettholde intracellulær homeostase ved å kontrollere konsentrasjonene av natrium- og kaliumioner. Dette vedlikeholdet av homeostase oppnås takket være:
Na + har en ekstracellulær konsentrasjon på ca. 142 mEq / L, mens dens intracellulære konsentrasjon bare er 12 mEq / L; K + er derimot mer konsentrert inne i cellen (140 mEq / L) enn utenfor den (4 mEq / L).
Selv om den elektriske ladningen til disse ionene ikke tillater passering gjennom membranen, er det ionekanaler som tillater det (selektivt), som fremmer bevegelse hvis kreftene som vanligvis beveger disse ionene også er tilstede..
Nå er disse forskjellene i konsentrasjon av stor betydning i bevaring av homeostase av organismen og må opprettholdes i en slags balanse som hvis tapt ville gi viktige organiske endringer.
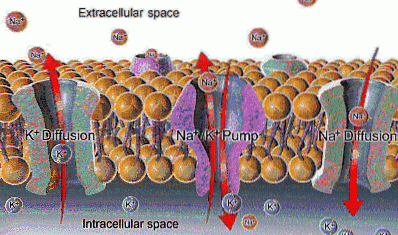
Funksjonen til Na + / K + -pumpen tillater ekstraksjon av natrium som har kommet inn ved diffusjon gjennom kanaler eller andre transportveier, og gjeninnføring av kalium som har diffundert ut, slik at bevaring av intra og ekstracellulære konsentrasjoner av disse ionene.
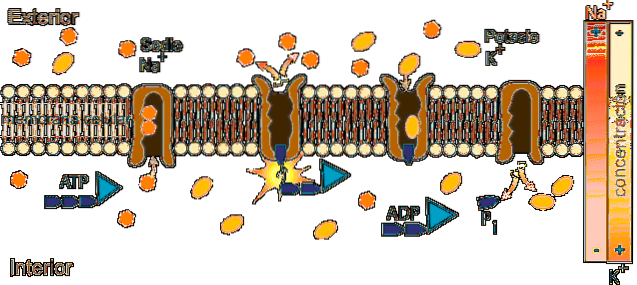
Virkningsmekanismen til ATPase Na + / K + består av en katalytisk syklus som involverer overføringsreaksjoner av en fosforylgruppe (Pi) og konformasjonsendringer av enzymet som går fra en E1-tilstand til en E2-tilstand og omvendt..
Operasjonen krever tilstedeværelse av ATP og Na + inne i cellen og K + i ekstracellulær væske.
Syklusen begynner i enzymets E1-konformasjonstilstand, der det er 3 cytosoliske bindingssteder til Na + og høy affinitet (Km på 0,6 mM) som er fullt opptatt fordi intraionkonsentrasjonen (12 mM) tillater.
I denne tilstanden (El) og med Na + festet til bindingsstedene, binder ATP seg til stedet i den cytosoliske sektoren av molekylet, hydrolyseres og en fosfatgruppe overføres til aspartat 376 og danner et høyenergi-acylfosfat som induserer en konformasjon endre til tilstand E2.
Konformasjonsendringen til E2-tilstanden innebærer at Na + -bindingsstedene passerer til utsiden, deres affinitet for ionet avtar sterkt og den frigjøres i det ekstracellulære væsken, mens affiniteten til K + -bindingsstedene samtidig øker og disse ionene festes på utsiden av pumpen.


Når Na + er frigjort og K + -bundet, oppstår hydrolysen av aspartylfosfat og konformasjonsendringen fra E2-tilstanden til E1-tilstanden blir tilbakeført, med gjeninnføring av de tomme Na + -bindingsstedene og de av okkupert K +.
Når denne endringen skjer, gjenoppretter stedene for Na + sin affinitet, og de for K + mister den, med hvilken K + frigjøres i cellen..
Na + / K + -pumpen er til stede i de fleste, om ikke alle, pattedyrceller, der det er av generell betydning ved å bidra til å opprettholde osmolariteten og dermed volumet.
Den kontinuerlige innføringen av natriumioner i cellen bestemmer en økning i det intracellulære antallet osmotisk aktive partikler, noe som induserer innføring av vann og økningen i volum som vil ende opp med å forårsake brudd på membranen og cellekollaps..
Ettersom disse pumpene bare introduserer 2 K + for hver 3 Na + de fjerner, oppfører de seg elektrisk, noe som betyr at de "dekompenserer" de interne elektriske ladningene, noe som favoriserer produksjonen av membranpotensialet som er karakteristisk for kroppens celler..
Dens betydning er også tydelig i forhold til cellene som utgjør opphissende vev, der handlingspotensialene er preget av inngangen til Na + -ionet, som depolariserer cellen, og utgangen av K +, som repolariserer den..
Disse ioniske bevegelsene er mulige takket være driften av Na + / K + -pumpene, som bidrar til produksjonen av de kjemiske gradientene som beveger de involverte ionene..
Uten at disse pumpene opererer i motsatt retning, vil konsentrasjonsgradientene til disse ionene forsvinne og den eksitatoriske aktiviteten forsvinne..
Et annet aspekt som fremhever den ekstreme betydningen av natrium-kaliumpumper, er relatert til nyrefunksjon, noe som ville være umulig uten dem.
Nyrefunksjon innebærer daglig filtrering av mer eller mindre 180 liter plasma og store mengder stoffer, hvorav noen må skilles ut, men mange må resorberes på nytt slik at de ikke går tapt gjennom urinen..
Reabsorpsjonen av natrium, vann og mange av de filtrerte stoffene avhenger av disse pumpene, som er plassert i den basolaterale membranen til cellene som utgjør epitelene til de forskjellige rørformede segmentene i nyrene..
Epitelcellene som strekker nyretubuli har ett ansikt som er i kontakt med tubulens lumen og kalles det apikale ansiktet, og et annet som er i kontakt med interstitiumet rundt tubuli og kalles basolateral..
Vannet og stoffene som blir absorbert, må først passere inn i cellen gjennom apikale og deretter inn i interstitium gjennom basolaterale.
Gjenopptaket av Na + er nøkkelen både i forhold til det, og i forhold til det til vann og andre stoffer som er avhengige av det. Den apikale innføringen av Na + i cellen krever at det er en gradient som beveger den, og det innebærer en veldig lav konsentrasjon av ionet inne i cellen..
Denne lave intracellulære konsentrasjonen av Na + produseres av natriumpumpene i den basolaterale membranen som jobber intensivt for å fjerne ionet fra cellene til interstitiet..

Ingen har kommentert denne artikkelen ennå.