De ketogenese Det er prosessen hvor man oppnår acetoacetat, β-hydroksybutyrat og aceton, som sammen kalles ketonlegemer. Denne komplekse og finregulerte mekanismen finner sted i mitokondriene, fra katabolismen av fettsyrer.
Å skaffe ketonlegemer finner sted når kroppen utsettes for uttømmende fasteperioder. Selv om disse metabolittene hovedsakelig er syntetisert i leverceller, er de funnet som en viktig energikilde i forskjellige vev, som skjelettmuskulatur og i hjerte- og hjernevev..
Β-hydroksybutyrat og acetoacetat er metabolitter som brukes som underlag i hjertemuskelen og hjernebarken. I hjernen blir ketonlegemer viktige energikilder når kroppen har tømt glukoselageret.
Artikkelindeks
Ketogenese betraktes som en veldig viktig fysiologisk funksjon eller metabolsk vei. Generelt foregår denne mekanismen i leveren, selv om det har vist seg at den kan utføres i andre vev som er i stand til å metabolisere fettsyrer..
Dannelsen av ketonlegemer er den viktigste metabolske avledningen av acetyl-CoA. Denne metabolitten er hentet fra den metabolske banen kjent som β-oksidasjon, som er nedbrytningen av fettsyrer.
Tilgjengeligheten av glukose i vevet der β-oksidasjon oppstår bestemmer den metabolske skjebnen til acetyl-CoA. I spesielle situasjoner er de oksyderte fettsyrene nesten helt rettet mot syntesen av ketonlegemer..
Hovedketonlegemet er acetoacetat eller aceteddiksyre, som hovedsakelig syntetiseres i leverceller. De andre molekylene som utgjør ketonlegemene er avledet fra acetoacetat.
Reduksjonen av aceteddiksyre gir opphav til D-β-hydroksybutyrat, det andre ketonlegemet. Aceton er en forbindelse som er vanskelig å nedbryte og produseres ved en spontan dekarboksyleringsreaksjon av acetoacetat (slik at den ikke krever inngrep av noe enzym), når den er tilstede i høye konsentrasjoner i blodet.
Betegnelsen på ketonlegemer er etablert ved konvensjon, siden β-hydroksybutyrat strengt tatt ikke har en ketonfunksjon. Disse tre molekylene er løselige i vann, noe som letter transporten av dem i blodet. Hovedfunksjonen er å gi energi til visse vev som skjelett- og hjertemuskulatur.
Enzymer involvert i dannelsen av ketonlegemer er hovedsakelig i lever- og nyreceller, noe som forklarer hvorfor disse to stedene er de viktigste produsentene av disse metabolittene. Dens syntese forekommer utelukkende i den mitokondrielle matrisen til celler.
Når disse molekylene er syntetisert, går de inn i blodstrømmen og går til vevet som krever dem, hvor de blir nedbrutt til acetyl-CoA..
Den metabolske skjebnen til acetyl-CoA fra β-oksidasjon avhenger av kroppens metabolske krav. Dette oksiderer til COto og HtoEller via sitronsyresyklusen eller syntesen av fettsyrer, hvis metabolismen av lipider og karbohydrater er stabil i kroppen.
Når kroppen trenger dannelse av karbohydrater, brukes oksaloacetat til fremstilling av glukose (glukoneogenese) i stedet for å starte sitronsyresyklusen. Dette skjer, som nevnt, når kroppen har en viss manglende evne til å oppnå glukose, i tilfeller som langvarig faste eller tilstedeværelse av diabetes.
På grunn av dette brukes acetyl-CoA som følge av oksidasjon av fettsyrer for produksjon av ketonlegemer..
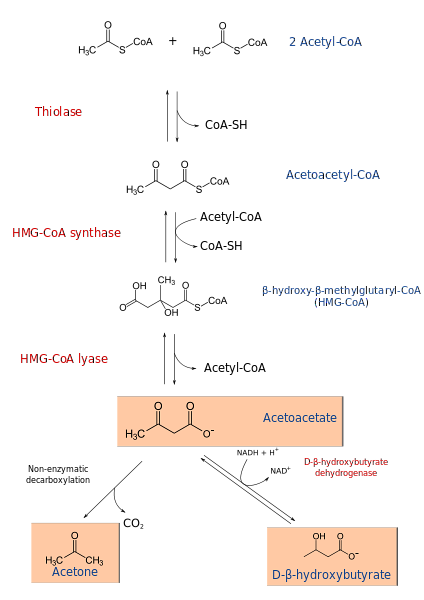
Ketogeneseprosessen begynner fra produktene av β-oksidasjon: acetacetyl-CoA eller acetyl-CoA. Når substratet er acetyl-CoA, består det første trinnet av kondensering av to molekyler, en reaksjon katalysert av acetyl-CoA transferase, for å produsere acetacetyl-CoA.
Acetacetyl-CoA kondenseres med en tredje acetyl-CoA gjennom virkningen av HMG-CoA-syntase for å produsere HMG-CoA (β-hydroksy-β-metylglutaryl-CoA). HMG-CoA nedbrytes til acetoacetat og acetyl-CoA ved virkning av HMG-CoA lyase. På denne måten oppnås det første ketonlegemet.
Acetoacetat reduseres til β-hydroksybutyrat ved intervensjon av β-hydroksybutyratdehydrogenase. Denne reaksjonen avhenger av NADH.
Hovedacetoacetatketonlegemet er en β-ketosyre, som gjennomgår ikke-enzymatisk dekarboksylering. Denne prosessen er enkel og produserer aceton og COto.
Denne reaksjonsserien gir altså ketonlegemer. Disse som er oppløselige i vann kan lett transporteres gjennom blodsirkulasjonen, uten behov for å forankres til en albuminstruktur, slik det er tilfellet med fettsyrer som er uoppløselige i vandig medium..
Fettsyremetabolisme produserer substratene for ketogenese, så disse to banene er funksjonelt relaterte.
Acetoacetyl-CoA er en hemmer av fettsyremetabolismen, siden det stopper aktiviteten til acyl-CoA dehydrogenase, som er det første enzymet av β-oksidasjon. I tillegg utøver den også inhibering av acetyl-CoA transferase og HMG-CoA syntase.
Enzymet HMG-CoA-syntase, underordnet av CPT-I (et enzym involvert i produksjonen av acylkarnitin i β-oksidasjon), spiller en viktig regulatorisk rolle i dannelsen av fettsyrer.
Fôring av organismer regulerer et komplekst sett med hormonsignaler. Karbohydratene, aminosyrene og lipidene som forbrukes i dietten, avsettes i form av triacylglyseroler i fettvev. Insulin, et anabole hormon, er involvert i syntesen av lipider og dannelsen av triacylglyseroler.
På mitokondrie-nivået kontrolleres β-oksidasjon ved inngang og deltakelse av noen substrater i mitokondriene. CPT I-enzymet syntetiserer Acyl Carnitine fra cytosolic Acyl CoA.
Når kroppen blir matet, blir Acetyl-CoA-karboksylase aktivert og citrat øker CPT I-nivåene, mens fosforyleringen avtar (reaksjon avhengig av syklisk AMP).
Dette forårsaker en akkumulering av malonyl CoA, som stimulerer syntesen av fettsyrer og blokkerer oksidasjonen deres, og forhindrer at det blir generert en meningsløs syklus..
I tilfelle av faste er karboksylaseaktiviteten veldig lav siden nivåene av CPT I-enzymet har blitt redusert, og det har også blitt fosforylert, noe som aktiverer og fremmer oksidasjon av lipider, som deretter vil tillate dannelse av ketonlegemer gjennom acetyl- CoA.
Ketonlegemer diffunderer ut av cellene der de ble syntetisert og transporteres til perifert vev i blodet. I disse vevene kan de oksyderes gjennom trikarboksylsyresyklusen.
I perifert vev oksyderes β-hydroksybutyrat til acetoacetat. Deretter aktiveres det tilstedeværende acetoacetatet ved virkningen av enzymet 3-ketoacyl-CoA transferase.
Succinyl-CoA fungerer som en CoA-giver ved å konvertere seg til succinat. Aktivering av acetoacetat skjer for å forhindre at succinyl-CoA konverterer til succinat i sitronsyresyklusen, med den koblede syntesen av GTP ved virkningen av succinyl-CoA-syntase.
Den resulterende acetoacetyl-CoA gjennomgår en tiolytisk nedbrytning og produserer to acetyl-CoA-molekyler som er innlemmet i trikarboksylsyresyklusen, bedre kjent som Krebs-syklusen..
Leverceller mangler 3-ketoacyl-CoA-transferase, og forhindrer at denne metabolitten aktiveres i disse cellene. På denne måten er det garantert at ketonlegemene ikke oksiderer i cellene der de ble produsert, men at de kan overføres til vevet der deres aktivitet er nødvendig..
I menneskekroppen kan høye konsentrasjoner av ketonlegemer i blodet forårsake spesielle forhold som kalles acidose og ketonemi..
Fremstillingen av disse metabolittene tilsvarer katabolismen av fettsyrer og karbohydrater. En av de vanligste årsakene til en patologisk ketogenesetilstand er den høye konsentrasjonen av eddikdikarbonatfragmenter som ikke blir nedbrutt av oksidasjonsveien til trikarboksylsyrer..
Som en konsekvens er det en økning i nivåene av ketonlegemer i blodet over 2 til 4 mg / 100 N og deres tilstedeværelse i urinen. Dette resulterer i forstyrrelse av den mellomliggende metabolismen av disse metabolittene..
Visse mangler i hypofysen neuroglandular faktorer som regulerer nedbrytning og syntese av ketonlegemer, sammen med lidelser i metabolismen av hydrokarboner, er årsaken til tilstanden til hyperketonemia..
Diabetes mellitus (type 1) er en endokrin sykdom som forårsaker økt produksjon av ketonlegemer. Utilstrekkelig insulinproduksjon deaktiverer transport av glukose til muskler, lever og fettvev, og akkumuleres dermed i blodet.
Celler i fravær av glukose begynner prosessen med glukoneogenese og nedbrytning av fett og protein for å gjenopprette metabolismen. Som en konsekvens reduseres oksaloacetatkonsentrasjonen og lipidoksidasjonen øker..
Det oppstår deretter en akkumulering av acetyl-CoA, som i fravær av oksaloacetat ikke kan følge sitronsyreveien, og dermed forårsake den høye produksjonen av ketonlegemer, karakteristisk for denne sykdommen..
Akkumuleringen av aceton oppdages ved tilstedeværelse i urinen og pusten til mennesker med denne tilstanden, og er faktisk et av symptomene som indikerer manifestasjonen av denne sykdommen.


Ingen har kommentert denne artikkelen ennå.