
De fibronektin Det er en klasse glykoprotein som tilhører den ekstracellulære matrisen. Generelt er denne typen proteiner ansvarlig for å feste eller binde cellemembranen til kollagenfibrene som finnes på utsiden.
Navnet "fibronektin" kommer fra et ord som består av to latinske ord, det første er "fiber" som betyr fiber eller filament, og det andre "Necter" som betyr å koble, koble, lime inn eller binde.
Fibronektin ble først visualisert i 1948 som en forurensning av fibrinogen fremstilt ved Cohns kalde etanolfraksjoneringsprosess. Dette ble identifisert som et unikt plasmaglykoprotein som hadde egenskapene til et kaldt uoppløselig globulin..
Dette proteinet har høy molekylvekt og har vært assosiert med en rekke funksjoner i vev. Disse inkluderer adhesjon mellom celle og celle, organisering av cytoskelettet, onkogen transformasjon, blant andre..
Fibronektin distribueres til mange steder i kroppen gjennom sin løselig form i blodplasma, cerebrospinalvæske, synovialvæske, fostervann, sædvæske, spytt og inflammatoriske ekssudater..
Forskere har rapportert at plasmakonsentrasjonen av fibronektin øker når gravide lider av svangerskapsforgiftning. Dermed har denne økningen i konsentrasjonen av fibronektin blitt innlemmet av spesialister for å diagnostisere denne tilstanden..
Artikkelindeks
Fibronektiner er store glykoproteiner, som har en molekylvekt på omtrent 440 kDa. De består av ca 2300 aminosyrer, som representerer 95% av proteinet, siden de andre 5% er karbohydrater.
De forskjellige analysene som har blitt utført på den genomiske og transkriptomiske sekvensen (messenger RNA) av proteinet, har indikert at det består av tre blokker med gjentatte homologe sekvenser, med lengder på 45, 60 og 90 aminosyrer hver..
De tre typer sekvenser utgjør mer enn 90% av den totale strukturen av fibronektiner. Homologe type I og II sekvenser er sløyfer knyttet til hverandre av disulfidbroer. Disse sløyfene inneholder henholdsvis 45 og 60 aminosyrerester..
Homologe type III-sekvenser tilsvarer 90 aminosyrer arrangert på en lineær måte og uten indre disulfidbroer. Imidlertid har noen av de indre aminosyrene av homologe type III-sekvenser frie sulfhydriske grupper (R-S-H).
De tre homologe sekvensene brettes og organiseres i en mer eller mindre lineær matrise for å danne to "dimeriske armer" av nesten identiske proteinunderenheter. Forskjeller mellom de to underenhetene oppstår fra modningshendelser etter transkripsjon.
Fibronektiner kan generelt sees på to måter. En åpen form som observeres når de avsettes på overflaten av membranen og som de er klare til å binde med en annen komponent i celleutvendig. Denne formen sees bare ved elektronmikroskopi..
Den andre formen kan sees i fysiologiske løsninger. Endene på hver arm eller forlengelse er brettet mot senteret av proteinet og forbinder seg gjennom karboksylendene på kollagenbindingsstedene. I denne formen har proteinet et kuleformet utseende.
Multi-adhesjonsegenskapene til fibronektin stammer fra tilstedeværelsen av forskjellige domener som har høye affinitetsverdier for forskjellige substrater og proteiner..
De "dimeriske armene" kan deles inn i 7 forskjellige funksjonelle domener. Disse er klassifisert i henhold til substratet eller domenet som hver enkelt binder seg til. For eksempel: Domene 1 og Domene 8 er fibrinproteinbindende domener.
Domene 2 har kollagenbindende egenskaper, domene 6 er en celleadhesjonsregion, det vil si at den lar den forankre seg på nesten hvilken som helst membran eller ytre overflate av celler. Funksjonene til domenene 3 og 5 er fortsatt ukjente i dag.
I domene 9 er karboksylenden eller C-terminale enden av proteinet lokalisert. Celleadhesjonsregionene i domene 6 har tripeptidet som består av aminosyresekvensen Arginin-Glycin-Asparagin (Arg-Gly-Asp).
Dette tripeptidet deles av flere proteiner som kollagen og integriner. Det er den minste strukturen som kreves for gjenkjenning av plasmamembranen av fibronektiner og integriner..
Fibronektin representerer en løselig og fri form i blodet når den er i kuleform. Imidlertid finnes det på celleoverflater og i den ekstracellulære matrisen i en ”åpen”, stiv og uoppløselig form..
Noen av prosessene der deltakelse av fibronektiner skiller seg ut er celle-til-celle-feste, cellebinding, forbindelse eller overholdelse av plasma- eller basalmembraner, stabilisering av blodpropp og sårheling..
Celler fester seg til et spesifikt sted på fibronektin gjennom et reseptorprotein kjent som "integrin." Dette proteinet krysser plasmamembranen til det indre av cellen.
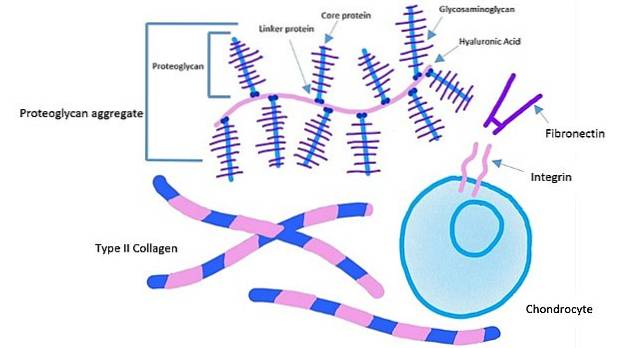
Det ekstracellulære domenet til integriner binder seg til fibronektin, mens det intracellulære domenet til integriner er festet til aktinfilamenter. Denne typen forankring gjør det mulig å overføre spenningen som genereres i den ekstracellulære matrisen til celleskjelettet..
Fibronektiner deltar i sårhelingsprosessen. Disse, i sin løselige form, avsettes på kollagenfibrene ved siden av såret, og hjelper migrasjonen av fagocytter, fibroblaster og celleproliferasjon i det åpne såret..
Selve helingsprosessen begynner når fibroblaster "spinner" fibronektin-nettverket. Dette nettverket fungerer som et slags stillas eller støtte for avsetning av de nye kollagenfibrene, heparansulfat, proteoglykan, chondrotin sultafo og de andre komponentene i den ekstracellulære matrisen som kreves for å reparere vevet..
Fibronektin er også involvert i bevegelsen av epidermale celler, da det gjennom det granulære vevet hjelper til med å omorganisere kjellermembranen som ligger under epidermis i vevet, noe som hjelper keratinisering å forekomme..
Alle fibronektiner har viktige funksjoner for alle celler; delta i prosesser så forskjellige som cellemigrasjon og differensiering, homeostase, sårheling, fagocytose, blant andre.



Ingen har kommentert denne artikkelen ennå.