De heterokromatin Det er en tettpakket del av kromatin (DNA og histonproteiner) av eukaryote kromosomer. Det er vanligvis assosiert med de "stille" områdene i genomet, det vil si med de som er transkripsjonelt inaktive.
Heitz, i 1928, var den første til å skille mellom to forskjellige typer kromatin på eukaryote kromosomer i mellomfasen, og beskrev eukromatin og heterochromatin på grunnlag av deres differensielle komprimering..
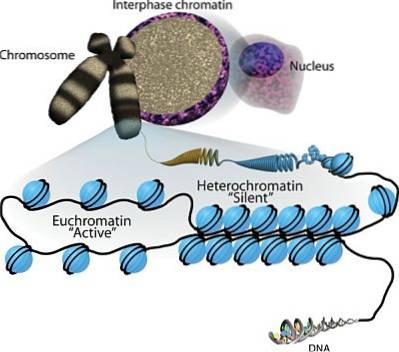
Hvis eukaryote kromosomer farges med forskjellige teknikker, spesifikke for DNA, avslører mikroskopiske observasjoner at det er områder av disse strukturene som flekker mer intenst enn andre. Disse regionene tilsvarer de hyperkompakte regionene til heterokromatin.
Heterokromatinisering av DNA, det vil si emballasjen, kan forekomme i en celle som respons på forskjellige faktorer og kan være fakultativ eller konstituerende.
Konstitutivt heterokromatin er et permanent trekk som generelt nedarves, mens fakultativ heterokromatin kan eller ikke kan være på et kromosom til enhver tid. Det beste eksemplet på konstituerende heterokromatin er det for en av de to X-kromosomene hos kvinner.
I eukaryoter "lagrer" og "komprimerer" heterokromatin de store genomene som kjennetegner dem, spesielt de regionene som består av repeterende sekvenser, gjenværende fraksjoner av invaderende retro-transposoner, transponerbare elementer, blant andre..
Artikkelindeks
Heterokromatin har ikke en veldig annen struktur enn kromatin som er mindre tettpakket, eukromatin.
For å forstå dette er det viktig å huske at eukaryote kromosomer består av et DNA-molekyl som er assosiert med proteiner som kalles histoner. Åtte histoner danner en oktamerisk kjerne kjent som et "nukleosom", rundt hvilket DNA brytes..
Assosiasjonen av DNA med histonproteiner oppstår takket være elektrostatisk interaksjon mellom de positive ladningene til de basiske restene av disse proteinene og de negative ladningene til fosfatgruppene i DNA-strengstrukturen..
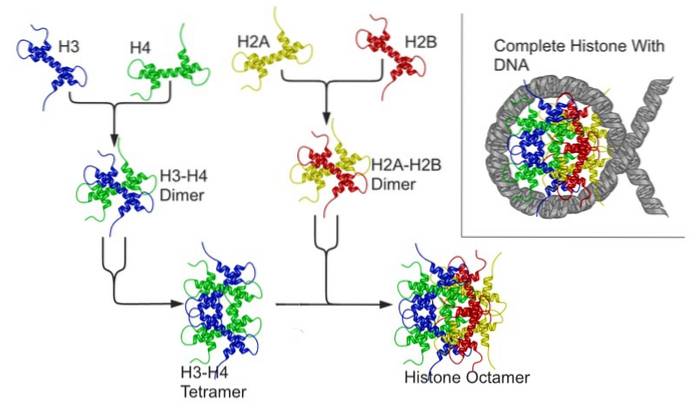
Hver histonoktamer er sammensatt av en tetramer av histonene H3 og H4, og av to dimerer av histonene H2A og H2B; omtrent hver kjerne av histoner rommer omtrent 146 basepar DNA.
Nukleosomer "kommer nærmere" hverandre takket være deltakelse av et annet histon, kjent som et kryss eller brohiston (lenker, på engelsk), som er histon H1.
Kromatin er deretter sammensatt av suksessive nukleosomer som komprimerer for å danne en fibrøs struktur med større tykkelse, men kortere i lengden..
Hvert histonprotein er preget av tilstedeværelsen av en aminosyre "hale" som kan gjennomgå kovalente enzymatiske modifikasjoner. Disse modifikasjonene har vist seg å påvirke graden av ekspresjon eller lyddemping av gener assosiert med nukleosomer, samt nivået på kromatinkomprimering..
Spesielt er heterokromatin karakterisert ved hypoacetylering av histoner i alle eukaryoter, og ved metylering av histon H3 ved lysinrest 9, bare for "høyere" eukaryoter..
Enzymer som er ansvarlige for å utføre disse modifikasjonene er kjent henholdsvis som histondeacetylaser og histonmetyltransferaser..
I tillegg til modifikasjoner i histoner, kan DNA også metyleres, noe som påvirker graden av komprimering av kromatin og tilsvarer den andre av de to epigenetiske mekanismene for organisering av det eukaryote genomet..
Heterokromatin, som kommentert i begynnelsen, kan være konstituerende eller fakultativ.
Konstitutivt heterokromatin er spesielt rikelig i genomiske regioner som har høy tetthet av repeterende sekvenser (for eksempel satellittelementer, for eksempel), der det er rikelig med tausede transponerbare elementer, i de sentromere regionene og i telomerer.
Det sies å være konstituerende fordi disse regionene i genomet forblir fortettet eller kompakte under celledeling. I en ikke-delende celle, derimot, er det meste av DNA eukromatisk, og det er bare noen få veldefinerte regioner av konstituerende heterokromatin.
Fakultativ heterokromatin er det som finnes i lokalene som er regulert i de forskjellige stadiene av utviklingen; så den representerer faktisk de "forbigående kondenserte" regionene som kan endres i henhold til cellulære signaler og genetisk aktivitet.
Siden heterokromatin er en viktig del av de telomere og sentromere områdene, utfører den transcendentale funksjoner fra synspunktet til celledeling og beskyttelse av kromosomale ender..
Sentromerene fungerer aktivt under celledeling, slik at dupliserte kromosomer beveger seg mot begge polene i delingscellen, mens resten av genene forblir inaktive og kompakte..
Komprimering av spesifikke regioner av eukaryote kromosomer er synonymt med genetisk demping, siden det faktum at heterokromatin er tettpakket, innebærer at transkripsjonsmaskineriet ikke er tilgjengelig for de underliggende gensekvensene.
Når det gjelder rekombinasjon, undertrykker heterokromatin denne prosessen og beskytter genomets integritet ved å forby "ulovlig" rekombinasjon mellom repeterende DNA-sekvenser spredt over hele genomet. Dette er spesielt viktig for kontroll av "parasittiske" transponerbare elementer, som blir tauset av heterokromatinisering..
Inntil for noen år siden ble det antatt at heterokromatisk DNA var et slags "søppel-DNA", siden forskere ikke fant en spesifikk funksjon for sekvensene som inngikk i disse regionene; husk at mer enn 80% av det genomiske DNA fra et menneske, for eksempel, ikke koder for cellulære proteiner eller for RNA-molekyler med regulatoriske funksjoner.
Imidlertid er det nå kjent at dannelsen av fakultativ heterokromatisk DNA er av største betydning for regulering av mange prosesser under utvikling og vekst av levende vesener, og at konstituerende heterokromatinregioner spiller en grunnleggende rolle fra et strukturelt synspunkt. av utsikten.
Det har blitt foreslått av mange forfattere at heterokromatin kan ha strukturelle funksjoner på eukaryote kromosomer. Denne påstanden er basert på det faktum at heterokromatiske regioner i et gitt kromosom skiller deler av det som har forskjellige mønstre for genetisk "aktivitet".
Med andre ord tjener de heterokromatiske regionene som "avstandsstykker" mellom forskjellige transkripsjonsaktive regioner, noe som kan være av stor betydning fra synspunktet til transkripsjonen av genene som er lokalisert der..


Ingen har kommentert denne artikkelen ennå.