De leukopoiesis Det er prosessen med dannelse og utvikling av leukocytter. Det er en del av hematopoiesis, prosessen der blodceller dannes, differensieres, utvikles og modnes, inkludert sammensetningen av erytrocytter, blodplater og leukocytter.
Hematopoiesis og derfor leukopoiesis er prosesser som finner sted i benmargen. I fosteret, i tillegg til benmargen, blir de også utført i lever og milt.
Fra fødselen til omtrent 20 år forekommer hematopoiesis i marg av alle bein. Fra 20-årsalderen blir marg av lange bein inaktiv, med unntak av den øvre delen av buksebenet og lårbenet. Den såkalte "røde margen", som er den aktive benmargen, har da ansvaret for hematopoiesis, for å skille den fra den gule, som er hematopoietisk inaktiv..
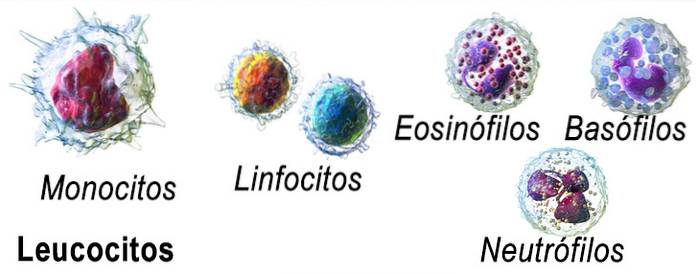
Leukopoiesis inkluderer differensiering, dannelse, utvikling og modning av forskjellige cellelinjer som gir opphav til fem typer celler:
- Neutrofile polymorfonukleære leukocytter eller granulocytter
- Polymorfonukleære eosinofiler
- Basofile polymorfonukleære celler
- Monocytter
- Noen lymfocytter.
Neutrofiler er de vanligste leukocyttene eller hvite blodcellene i blodet. Til tross for at det er 500 ganger mer erytrocytter i omløp enn leukocytter, er 75% av cellene i benmargen av den myeloide serien som produserer leukocytter.
Artikkelindeks
I benmargen er det celler som kalles "stamceller" eller "stamceller", stamceller eller "hemocytoblaster". Dette er stamceller for alle blodceller fra benmargen, men de gir også opphav til osteoklaster, Kupffer-celler, mastceller, dendritiske celler og Langerhans-celler..
Det som skjer først i leukopoieseprosessen er at disse stamcellene deler seg og gir opphav til celler som kalles "kompromitterte stamceller", som er myeloide stamceller og lymfoide stamceller..
I sin tur skiller myeloide celler seg og produserer røde blodceller eller erytrocytter, blodplater, granulocytter eller polymorfonukleære celler og monocytter. Lymfoide celler vil gi opphav til lymfocytter.
For leukopoiesis-prosessen er utviklingen av monocytter og granulocytter beskrevet. Så, myeloide celler skiller seg ut i monoblaster og myeloblaster. Monoblaster gir opphav til "promonocytter", en prosess der en endring skjer i formen til kjernen, som begynner å bøye seg. Promonocytter blir monocytter. På dette utviklingsstadiet får kjernen sin endelige hesteskoform..
Myeloblaster gir opphav til tre cellelinjer: basofile promyelocytter, eosinofile promyelocytter og nøytrofile promyelocytter. De er celler med cytoplasmatiske granuler som flekker med fargestoffer med forskjellig pH.
Promyelocytter gir opphav til myelocytter, og danner dermed basofile myelocytter, eosinofile myelocytter og nøytrofile myelocytter. I disse cellene begynner kjernene å endre form.
Deretter får kjernen til disse cellene en "U" -form og de "metamyelocytter" eller båndcellene, nøytrofile, basofile og eosinofile celler dannes..
Basofile båndceller utvikler seg ferdig ved å trekke sammen kjernen for å danne en "S" -formet kjerne og bli basofiler..
Eosinofile båndceller danner en bilobed kjerne og gir opphav til eosinofiler og nøytrofile båndceller utvikler en polylobulert kjerne og danner nøytrofiler.
Stamceller forpliktet til lymfoide avstamning eller lymfoide stamceller gir opphav til lymfoblaster. Disse cellene skiller seg igjen og danner såkalte "prolymfocytter".
Prolymfocytter fortsetter å utvikle seg for å gi opphav til lymfocytter. To typer lymfocytter dannes i benmargen: B-lymfocytter og T-lymfocytter. B-lymfocytter er aktive celler. De forlater benmargen i blodet og derfra kan de gå til lymfeknuter. Disse cellene er modne og aktive celler.
T-lymfocytter produsert i beinmarg er umodne celler som passerer inn i blodet og når thymus eller lymfeknuter eller andre lymfoide organer der modning eller aktiveringsprosess slutter..
Spredning og differensiering av stamceller og de forskjellige stamcellene som er involvert inntil dannelsen av leukocytter skyldes en rekke hormonelle faktorer, som virker spesifikt i de forskjellige differensieringsstadiene av leukopoies.
Interleukiner (IL) og kolonistimulerende faktorer (CSF) er de viktigste stimulatorene for stamcelledifferensiering og påfølgende spredning og modning av forskjellige leukocyttcellelinjer.
I nærvær av interleukin 3 og 5 (IL3 Y 5) og agranulocyttkolonistimulerende faktor (aG-CSF), differensieres stamceller til monoblaster. Dannelsen av myeloblaster avhenger av tilstedeværelsen av IL3, IL5 og den granulocyttkolonistimulerende faktoren (G-CSF).
Interleukin 4 (IL4) deltar i differensieringen av myeloblaster langs basofil linjen. Andre faktorer er beskrevet som granulocytt- og makrofagkolonistimulerende faktor (GM-CSF) og makrofagkolonistimulerende faktor (M-CSF)..
Noen studier viser at fraværet av noen faktorer, i visse tilfeller, kan erstattes av resten, dette innebærer felles deltakelse av flere faktorer.
Hvite blodlegemer, spesielt nøytrofiler, har en veldig kort halveringstid. Sirkulerende granulocytter har en gjennomsnittlig halveringstid på 4 til 8 timer, hvoretter de må byttes ut. Halveringstiden i vev er 4 til 5 dager.
Monocytter i blod har en halveringstid på 10 til 20 timer, og når de går over i vev og blir makrofager, kan de vare i flere måneder. Lymfocytter lever i flere uker eller måneder og fortsetter å sirkulere mellom blod og lymfe.
Disse egenskapene krever et signalsystem for erstatning og økt produksjon når infeksjoner oppstår som krever "ekstra" mengder leukocytter. Sammen er disse mekanismene som opprettholder produksjon og frigjøring etter behov det som kalles "regulatorisk prosess for leukopoiesis"..
Reguleringen av differensiering og produksjon av leukocytter avhenger av en serie stoffer, blant hvilke regulatoriske faktorer (vekstfaktorer) som er glykoproteiner eller hormoner som stimulerer differensiering av stamceller og som også holder sirkulerende celler aktive..
Når leukocytter dannes i benmargen, slippes ikke alle ut i blodet, en del forblir i margen som en reserve til sirkulasjonssystemet krever det. Antall granulocytter som er lagret i reserve i benmargen tredobler antall sirkulerende leukocytter.
Disse reservene tillater levering i omtrent 5 eller 6 dager. Hvis det oppstår en smittsom eller inflammatorisk prosess, frigjør makrofager og aktiverte T-lymfocytter faktorer som stimulerer en økning i leukocyttdannelse, og øker kolonistimulerende faktorer..
Dermed forekommer leukocytose (økning i leukocytter i blodet) som følger med noen smittsomme prosesser. Hos mus og sannsynligvis hos mennesker er proteiner dannet av genet involvert i regulering av spredning og fornyelse av stamceller i benmargen. scl (leukemi i stamceller).


Ingen har kommentert denne artikkelen ennå.