De ontogeni det er prosessen der utviklingen av et individ skjer. Fenomenet begynner med befruktning, og strekker seg til aldring av organiske vesener. Feltet biologi med ansvar for å studere ontogeni er utviklingsbiologi.
I denne prosessen forekommer "oversettelsen" av genotypen - all den genetiske informasjonen til en biologisk enhet - til fenotypen vi kan observere. Den mest dramatiske transformasjonen skjer i de tidlige stadiene av utviklingen, med transformasjonen av en celle til et komplett individ.
I dag er sammensmeltingen av utviklingsbiologi og evolusjonsteori, kjent som evo-devo, en veldig populær kunnskap som vokser med store sprang. Dette nye feltet tar sikte på å forklare utviklingen av det enorme mangfoldet av morfologier som levende organismer viser..
Artikkelindeks
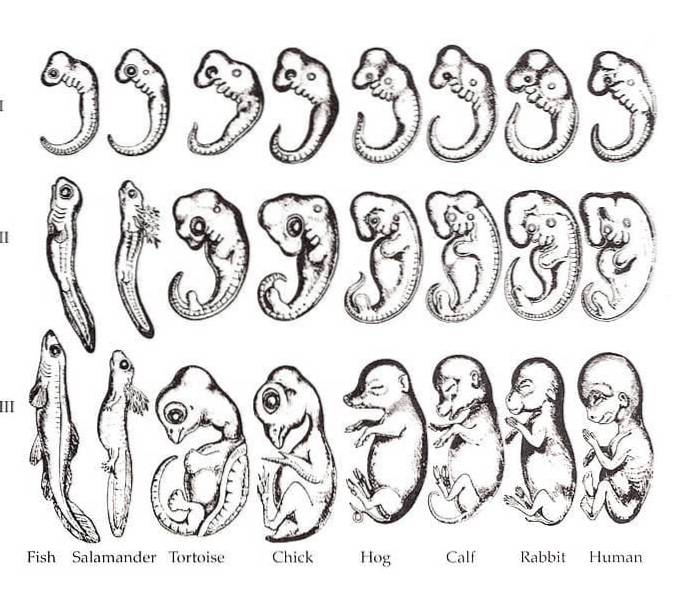
Forholdet mellom ontogeni og fylogeni var et dominerende syn gjennom det 21. århundre. Det er allment kjent at forskjellige arter av organismer er mye mer lik hverandre i sine embryonale stadier enn i voksne former. I 1828 la Karl Ernst von Baer merke til dette mønsteret i sybphylum Vertebrata.
Baer bemerket at det i forskjellige arter av tetrapoder er visse likheter i embryoet, som gjeller, notokord, segmentering og finneformede lemmer..
Disse dannes før de typiske egenskapene som gjør at gruppen det er snakk om å få diagnosen i en mer spesifikk hierarkisk klassifikasjonsrekkefølge.
Denne ideen ble omformulert av den berømte - og en av Charles Darwins mest lidenskapelige tilhengere - tyskfødte biolog Ernst Haeckel..
Haeckel er kreditert den berømte frasen "ontogeny recapitulates fylogeny." Med andre ord foreslår rekapitulering at utviklingen av en organisme gjentar dens evolusjonære historie fra de voksne formene til sine forfedre..
Selv om uttrykket er kjent i dag, var det på midten av det 21. århundre klart at Haeckels forslag sjelden blir oppfylt..
S. J. Gould, den berømte paleontologen og evolusjonsbiologen, la fram sine ideer om rekapitulering i det han kalte "terminaladdisjonsprinsippet." For Gould kan rekapitulering skje så lenge evolusjonær endring skjer ved suksessiv tilsetning av stadier på slutten av en forfedres ontogeni..
På samme måte måtte det også oppfylles at den tidsmessige varigheten av forfedres ontogeni måtte forkortes etter hvert som avstammen utviklet seg.
I dag har moderne metoder klart å motbevise begrepet addisjon foreslått av den biogenetiske loven.
For Haeckel skjedde dette tillegget på grunn av den kontinuerlige bruken som ble gitt til organene. Imidlertid har de evolusjonære implikasjonene av bruk og bruk av organer blitt forkastet.
Det er for tiden kjent at gjellebuene i embryonale stadier av pattedyr og reptiler aldri har form som tilsvarer voksen fisk.
I tillegg er det variasjoner i tidspunktet eller tidspunktet for visse utviklingstrinn. I evolusjonær biologi kalles denne endringen heterokroni..
Ontogeny omfatter alle prosesser for utvikling av organiske vesener, som begynner med befruktning og slutter med aldring..
Logisk sett forekommer de mest dramatiske transformasjonene i de første trinnene, hvor en enkelt celle er i stand til å danne et helt individ. Deretter vil vi beskrive ontogeniprosessen, med vekt på embryonale stadier.
Under oogeneseprosessen forbereder et egg (den kvinnelige kjønnscellen, også kalt et egg) for befruktning og de tidlige stadiene av utviklingen. Dette skjer ved å samle reservemateriale for fremtiden..
Cytoplasmaet i egget er et miljø rikt på forskjellige biomolekyler, hovedsakelig messenger-RNA, ribosomer, overførings-RNA og andre maskiner som er nødvendige for syntese av proteiner. Cellekjernen gjennomgår også betydelig vekst.
Sæd krever ikke denne prosessen, deres strategi er å eliminere så mye cytoplasma som mulig og kondensere kjernen for å bevare små dimensjoner..
Arrangementet som markerer begynnelsen på ontogeni er befruktning, som involverer foreningen av en mannlig og kvinnelig gamet, generelt under seksuell reproduksjon..
I tilfelle ekstern befruktning, som forekommer i mange marine organismer, blir begge kjønnsceller utvist i vannet og blir funnet tilfeldig.
I befruktning integreres det diploide antallet til individet på nytt og tillater kombinasjonsprosesser mellom fader- og morsgenene.
I visse tilfeller er sæd ikke nødvendig for å aktivere utvikling. Men hos de fleste individer utvikler embryoet seg ikke på riktig måte. På samme måte kan noen arter reprodusere ved partenogenese, der normal embryoutvikling skjer uten behov for sæd..
Derimot krever noen egg aktivering av sædceller, men innlemmer ikke genetisk materiale til denne mannlige kjønnscellen i embryoet..
Sæd og egg må gjenkjennes riktig slik at alle hendelser etter befruktning kan finne sted. Denne anerkjennelsen formidles av en serie artsspesifikke proteiner. Det er også barrierer som forhindrer at et egg når det er befruktet, nås av en andre sædceller.
Etter befruktning og aktivering av egget, oppstår de første utviklingsstadiene. I segmentering deler embryoet seg gjentatte ganger for å bli en gruppe celler kalt blastomerer..
I løpet av denne siste perioden forekommer ingen cellevekst, bare underoppdelingen av massen finner sted. Til slutt er det hundrevis eller tusenvis av celler som gir vei til blastula-staten.
Når embryoet utvikler seg, får det en polaritet. Av denne grunn er det mulig å skille mellom plantepolen, som ligger i den ene enden, og dyrepolen, rik på cytoplasma. Denne aksen gir et referansepunkt for utvikling.
Avhengig av hvor mye eggeplomme egget har, og fordelingen av stoffet, kan egget klassifiseres som oligolecytter, heterolecytter, telolecytter og sentrolecytter..
Førstnevnte har, som navnet antyder, en liten mengde eggeplomme og den er mer eller mindre jevnt fordelt over egget. Vanligvis er størrelsen liten. Heterolecytter har mer eggeplomme enn oligolecytter, og eggeplomme er konsentrert i den vegetative polen.
Telolecitos har en rikelig mengde eggeplomme, og opptar nesten hele egget. Til slutt har centrolecitos hele eggeplommen konsentrert i den sentrale regionen av egget..
Blastula er en masse celler. Hos pattedyr kalles denne cellegruppen en blastocyst, mens cellene i de fleste dyr er ordnet rundt et sentralt væskehulrom, kalt blastocele..
I blastula-tilstand har det vært mulig å vise en stor økning når det gjelder mengden DNA. Imidlertid er størrelsen på hele embryoet ikke mye større enn den opprinnelige zygoten..
Gastrulering omdanner den sfæriske og enkle blastulaen til en mye mer kompleks struktur med to bakterielag. Denne prosessen er heterogen hvis vi sammenligner de forskjellige linjene til dyr. I noen tilfeller dannes et andre lag uten å lage et indre hulrom.
Åpningen til tarmen kalles blastopore. Blastopores skjebne er en veldig viktig egenskap for delingen av to store linjer: protostomates og deuterostomes. I den første gruppen gir blastopore munnen, mens i den andre blastopore gir anus.
Dermed har gastrula to lag: et ytre lag som omgir blastocele, kalt ektoderm, og et indre lag kalt endoderm..
De fleste dyr har et tredje bakterielag, mesoderm, som ligger mellom de to lagene nevnt ovenfor. Mesoderm kan dannes på to måter: Cellene oppstår fra en ventral region av blastopore-leppen, og derfra sprer de seg eller oppstår fra den sentrale regionen av archnteron-veggene..
På slutten av gastrulering dekker ektoderm embryoet og mesoderm og endoderm ligger i den indre delen. Med andre ord har cellene en annen sluttposisjon enn den de startet i..
Coelom er et kroppshulrom som er omgitt av mesoderm. Dette skjer fordi blastocele er nesten fullstendig fylt med mesoderm under gastruleringsprosessen..
Dette coelomatiske hulrommet kan vises på to måter: schizocelic eller enterocelic. Imidlertid er begge coeloms funksjonelt likeverdige.
Organogenese består av en serie prosesser der hvert av organene dannes.
De mest relevante hendelsene inkluderer migrering av bestemte celler til stedet der de er nødvendige for å danne nevnte organ..
Under utvikling er det bestemt at epigenesen fortsetter i tre trinn: mønsterdannelse, bestemmelse av kroppsposisjon og induksjon av riktig posisjon for ekstremiteter og forskjellige organer.
For å generere en respons er det visse genprodukter, kalt morfogener (definisjonen av disse enhetene er teoretisk, ikke kjemisk). Disse fungerer takket være dannelsen av en differensiell gradient som gir romlig informasjon.
Når det gjelder gener involvert, spiller homeotiske gener en grunnleggende rolle i utviklingen av individer, siden de definerer identiteten til segmentene.


Ingen har kommentert denne artikkelen ennå.