
EN operon består av en gruppe sekvensielt ordnede gener som regulerer hverandre, som koder for proteiner som er funksjonelt relaterte, og som finnes i hele genomet av bakterier og de "forfedre" genomene.
Denne reguleringsmekanismen ble beskrevet av F. Jacob og J. Monod i 1961, et faktum som ga dem Nobelprisen i fysiologi og medisin i 1965. Disse forskerne foreslo og demonstrerte operasjonen av operoner gjennom gener som koder for enzymene som kreves av Escherichia coli for bruk av laktose.

Operoner har ansvaret for å koordinere proteinsyntese i henhold til behovene til hver celle, det vil si at de bare uttrykkes for å generere proteiner på det tidspunktet og på det nøyaktige stedet der de er nødvendige..
Genene som finnes i operoner er generelt strukturelle gener, noe som betyr at de koder for viktige enzymer som er direkte involvert i metabolske veier i cellen. Disse kan være syntesen av aminosyrer, energi i form av ATP, karbohydrater, etc..
Operoner finnes også ofte i eukaryote organismer, men i motsetning til prokaryote organismer transkriberes ikke regionen av operon i eukaryoter som et enkelt messenger RNA-molekyl..
Artikkelindeks
Det første viktige fremskrittet angående operoner laget av François Jacob og Jacques Monod var å fokusere på problemet med "enzymatisk tilpasning", som bare besto i utseendet til et spesifikt enzym når cellen var i nærvær av et substrat.
En slik respons fra celler på underlag hadde blitt observert i bakterier i mange år. Forskerne lurte imidlertid på hvordan cellen bestemte nøyaktig hvilket enzym den måtte syntetisere for å metabolisere det substratet..
Jacob og Monod observerte at bakterieceller, i nærvær av galaktoselignende karbohydrater, produserte 100 ganger mer β-galaktosidase enn under normale forhold. Dette enzymet er ansvarlig for å bryte ned β-galaktosider slik at cellen bruker dem metabolisk.
Dermed kalte begge forskere galaktosidtypen karbohydrater som "induktorer", siden de var ansvarlige for å indusere en økning i syntesen av β-galaktosidase..
Likeledes fant Jacob og Monod en genetisk region med tre gener som ble kontrollert på en koordinert måte: Z-genet, som koder for β-galaktosidase-enzymet; Y-genet, som koder for enzymet laktosepermease (galaktosidtransport); og gen A, som koder for enzymet transacetylase, som også er viktig for assimilering av galaktosider.
Gjennom påfølgende genetiske analyser avklarte Jacob og Monod alle aspekter av den genetiske kontrollen av laktoseoperonen, og konkluderte med at segmentet av genene Z, Y og A utgjør en enkelt genetisk enhet med koordinert uttrykk, som var det de definerte som "operon".
Operonmodellen ble først beskrevet nøyaktig i 1965 av Jacob og Monod for å forklare reguleringen av gener som er transkribert og oversatt for enzymene som kreves i Escherichia coli for å metabolisere laktose som energikilde.
Disse forskerne foreslo at transkripsjonene av genet eller settet med gener som er lokalisert fortløpende blir regulert av to elementer: 1) et regulatorisk gen eller repressorgenet 2) og et operatørgen eller operatorsekvens.
Operatørgenet finnes alltid ved siden av strukturgenet (-ene) hvis uttrykk er ansvarlig for regulering, mens repressorgenet koder for et protein kalt "repressor" som binder seg til operatøren og forhindrer transkripsjon..
Transkripsjon blir undertrykt når repressoren er koblet til operatorgenet. På denne måten uttrykkes ikke den genetiske ekspresjonen av genene som koder for enzymene som er nødvendige for å assimilere laktose, og kan derfor ikke metabolisere disakkaridet..
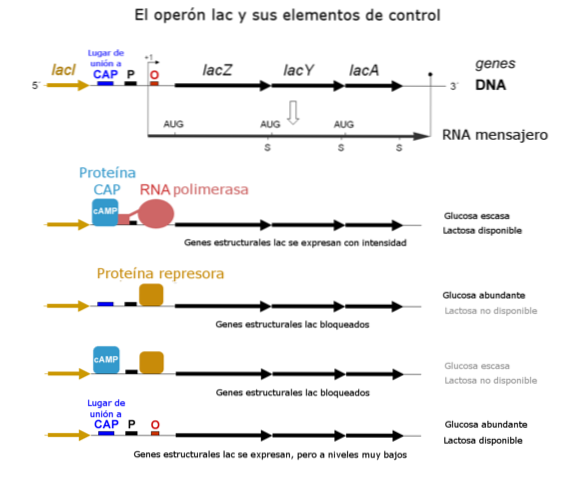
For tiden er det kjent at bindingen av repressoren til operatøren forhindrer, med steriske mekanismer, at RNA-polymerase binder seg til promoterstedet slik at den begynner å transkribere gener.
Promotorsiden er "stedet" som RNA-polymerase gjenkjenner for å binde og transkribere gener. Ettersom den ikke kan binde, kan den ikke transkribere noen av genene i sekvensen.
Operatorgenet ligger mellom en genetisk region i sekvensen kjent som promoter og strukturgenene. Imidlertid identifiserte ikke Jacob og Monod denne regionen i sin tid.
Det er for tiden kjent at den komplette sekvensen som inkluderer strukturgenet eller -genene, operatøren og promoteren, egentlig er det som utgjør et "operon"..
Operoner er klassifisert i bare tre forskjellige kategorier som avhenger av måten de reguleres på, det vil si at noen uttrykkes kontinuerlig (konstituerende), andre trenger noe spesifikt molekyl eller faktor for å aktivere (induserbar) og andre uttrykkes kontinuerlig til at induseren er uttrykt (undertrykkelig).
De tre typer operoner er:
Operoner av denne typen reguleres av molekyler i miljøet som aminosyrer, sukker, metabolitter, etc. Disse molekylene er kjent som induktorer. Hvis molekylet som fungerer som en induserer ikke blir funnet, transkriberes ikke operongenene aktivt.
I induserbare operoner binder den frie repressoren til operatøren og forhindrer transkripsjon av genene som finnes i operonet. Når induseren binder seg til repressoren, dannes et kompleks som ikke kan binde seg til repressoren, og derfor blir genene til operonen oversatt.
Disse operonene er avhengige av spesifikke molekyler: blant andre aminosyrer, sukker, kofaktorer eller transkripsjonsfaktorer. Disse er kjent som corepressors og fungerer på en helt motsatt måte til induktorer..
Først når corepressoren binder seg til repressoren, stopper transkripsjonen, og transkripsjonen av genene som finnes i operonet skjer derfor ikke. Da stopper transkripsjonen av et undertrykkbart operon bare med tilstedeværelsen av corepressoren.
Denne typen operoner er ikke regulert. De transkriberes konstant aktivt, og i tilfelle en mutasjon som påvirker sekvensen til disse genene, kan levetiden til cellene som inneholder dem påvirkes og generelt utløse programmert celledød..
Det tidligste og mest anerkjente eksemplet på funksjonen til en operon er Operon lac (laktose). Dette systemet er ansvarlig for å transformere laktose, et disakkarid, til monosakkaridene glukose og galaktose. Tre enzymer virker i denne prosessen:
- Β-galaktosidase, ansvarlig for å omdanne laktose til glukose og galaktose.
- Laktosepermease, ansvarlig for transport av laktose fra det ekstracellulære mediet til det indre av cellen og
- Transketylase, som tilhører systemet, men har en ukjent funksjon
Operonen trp (tryptofan) av Escherichia coli kontrollerer syntesen av tryptofan, med korisminsyre som forløper. Innenfor dette operonet er gener for fem proteiner som brukes til produksjon av tre enzymer:
- Det første enzymet, kodet av genene E og D, katalyserer de to første reaksjonene av tryptofanveien og er kjent som antranilatsyntetase.
- Det andre enzymet er glyserolfosfat og det katalyserer nedstrøms trinn av antranilatsyntetase.
- Det tredje og siste enzymet er tryptofansyntetase, som er ansvarlig for å produsere tryptofan fra indol-glyserolfosfat og serin (dette enzymet er et produkt av gener B og A)
Ingen har kommentert denne artikkelen ennå.