
De dihybridisme, I genetikk definerer den samtidig studie av to forskjellige arvelige karakterer, og i forlengelse, av de hvis manifestasjon avhenger av to forskjellige gener, selv om det har samme karakter.
De syv trekkene som Mendel analyserte, var nyttige for ham når han la fram sin teori om arv av tegn fordi blant annet gener som var ansvarlige for deres manifestasjon hadde kontrasterende alleler hvis fenotype var lett å analysere, og fordi hver enkelt bestemte uttrykket enkelt karakter.
Det vil si at de var monogene egenskaper hvis hybridtilstand (monohybrider) tillot å bestemme dominans / recessivitetsforholdet mellom allelene til det ene genet..
Da Mendel analyserte den felles arven til to forskjellige karakterer, fortsatte han som han hadde med enkeltkarakterene. Han fikk dobbel hybrider (dihybrider) som tillot ham å sjekke:
Vi vet nå at arven til tegn er litt mer kompleks enn hva Mendel observerte, men også at Mendel i sin grunnleggende retning var helt korrekt..
Den påfølgende utviklingen av genetikk gjorde det mulig å demonstrere at dihybridkryss og deres analyse (dihybridisme), slik Bateson opprinnelig var i stand til å demonstrere, kunne være en uuttømmelig kilde til funn i denne kraftige og fremvoksende vitenskapen fra det 20. århundre..
Takket være deres smarte bruk klarte de å gi genetikeren en noe klarere ide om genenes oppførsel og natur..
Artikkelindeks
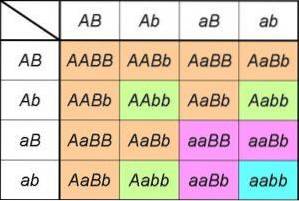
Hvis vi analyserer produktene fra et monohybridkors Aa X Aa, vi kan merke at det er lik å utvikle det bemerkelsesverdige produktet (TIL+til)to= AA + 2Aa + aa.
Uttrykket til venstre inkluderer de to typene gameter som en av foreldrene som er heterozygote for genet, kan produsere TIL/til; ved å kvadrere indikerer vi at begge foreldrene har samme konstitusjon for genet som studeres [det vil si at det er et monohybridkors (TIL+til) X (TIL+til)].
Uttrykket til høyre gir oss genotypene (og derfor blir fenotypene utledet) og forventede proporsjoner avledet fra krysset.
Derfor kan vi direkte observere de genotypiske proporsjonene avledet fra den første loven (1: 2: 1), så vel som de fenotypiske proporsjonene forklart av den (1AA+toAa= 3TIL_ for hver 1aa, eller fenotypisk forhold 3: 1).
Hvis vi nå vurderer et kryss for å analysere arven til et gen B, uttrykkene og proporsjonene vil være de samme; faktisk vil det være slik for ethvert gen. I et dihybridkors har vi derfor utviklingen av produktene fra (TIL+til)to X (B+b)to.
Eller hva er det samme, hvis dihybridkorset involverer to gener som deltar i arven av to ikke-relaterte tegn, vil de fenotypiske proporsjonene være de som er forutsagt av den andre loven: (3TIL_:1aa) X (3B_:1baby) = 9TIL_B_: 3TIL_baby: 3aaB_:1aabb).
Disse er selvfølgelig avledet fra de ordnede genotypeforholdene 4: 2: 2: 2: 2: 1: 1: 1: 1 som er et resultat av produktet fra (TIL+til)to X (B+b)to= (AA + 2Aa + aa) X (BB + toBb + baby).
Vi inviterer deg til å se selv for nå å analysere hva som skjer når de fenotypiske forholdene 9: 3: 3: 1 i et dihybridkors "avviker" fra disse klare og forutsigbare matematiske forhold som forklarer den uavhengige arven til to kodede tegn. Av forskjellige gener.
Det er to hovedmåter som dihybridkryss avviker fra det som er "forventet". Den første er den der vi analyserer felles arv av to forskjellige tegn, men de fenotypiske proporsjonene observert i avkommet gir en klar overvekt til manifestasjonen av foreldrenes fenotyper..
Mest sannsynlig er det et tilfelle av koblede gener. Det vil si at de to gener som er under analyse, selv om de er på forskjellige steder, er så nær hverandre fysisk at de har en tendens til å bli arvet sammen, og åpenbart blir de ikke distribuert uavhengig..
Den andre omstendigheten, som også er ganske vanlig, kommer av det faktum at en liten minoritet av arvelige egenskaper er monogene..
Tvert imot, mer enn to gener deltar i manifestasjonen av de fleste arvelige egenskaper..
Av denne grunn er det alltid mulig at de genetiske interaksjonene som er etablert mellom genene som deltar i manifestasjonen av en enkelt karakter, er komplekse og går utover et enkelt forhold mellom dominans eller recessivitet som observert i relasjonene alleliske som er typiske for monogene egenskaper..
For eksempel, i manifestasjonen av et trekk, kan omtrent fire enzymer delta i en bestemt rekkefølge for å gi opphav til sluttproduktet som er ansvarlig for den fenotypiske manifestasjonen av den ville fenotypen..
Analysen som gjør det mulig å identifisere antall gener fra forskjellige loci som deltar i manifestasjonen av et genetisk trekk, samt rekkefølgen de virker i, kalles epistasisanalyse og er kanskje den som oftest definerer hva vi kaller genetisk analyse. i sin mest klassiske forstand.
På slutten av dette innlegget presenteres de fenotypiske proporsjonene som er observert i de vanligste tilfellene av epistase - og dette tar bare hensyn til dihybridkryss..
Ved å øke antall gener som deltar i manifestasjonen av samme trekk, øker åpenbart kompleksiteten av geninteraksjoner og deres tolkning..
I tillegg, som igjen kan tas som den gyldne regel for en korrekt diagnose av epistatiske interaksjoner, kan utseendet på nye fenotyper som ikke er tilstede i foreldregenerasjonen, verifiseres..
Til slutt, bortsett fra å tillate analyse av utseendet til nye fenotyper og deres andel, tillater analysen av epistase også å bestemme den hierarkiske rekkefølgen som de forskjellige gener og deres produkter må manifestere seg i en gitt vei for å redegjøre for fenotypen som er knyttet til dem..
Det mest grunnleggende eller tidlige manifestasjonsgenet er epistatisk over alle de andre, siden uten produkt eller handling, for eksempel, vil ikke de nedstrøms det være i stand til å uttrykke seg, noe som derfor vil være hypostatisk for det..
Et gen / produkt på tredjeplass i hierarkiet vil være hypostatisk for de to første, og epistatisk for alle andre som er igjen i denne genuttrykkelsesveien..



Ingen har kommentert denne artikkelen ennå.