
De pachytene eller paquinema er den tredje fasen av meiotisk profase I; i den bekreftes rekombinasjonsprosessen. I mitose er det en profase, og i meiose to: profase I og profase II.
Tidligere, bortsett fra profase II, ble kromosomene duplisert, hver med opphav til en søsterkromatid. Men bare i profase parrer jeg homologer (duplikater) og danner bivalenter.

Begrepet paquiteno kommer fra gresk og betyr "tykke tråder". Disse "tykke trådene" er de sammenkoblede homologe kromosomene som etter duplisering danner tetrader. Det vil si fire "tråder" eller strenger som får hvert kromosom til å se tykkere.
Det er unike aspekter ved meiotisk profase I som forklarer de iboende egenskapene til pachyten. Bare i meiose-profase I pachytene rekombineres kromosomer.
For å gjøre dette bekreftes anerkjennelsen og matching av homologene. Som i mitose, må det være duplisering av kromatidene. Men det er bare i meiosen I pachytene det dannes båndutvekslingskomplekser, som vi kaller chiasmata..
I dem forekommer det som definerer rekombinasjonskraften til meiose: krysset mellom kromatider av homologe kromosomer.
Hele prosessen med DNA-utveksling er mulig takket være det tidligere synaptonemiske komplekset. Dette multiproteinkomplekset gjør at homologe kromosomer kan pares (synaps) og rekombineres..
Artikkelindeks
Det synaptonemiske komplekset (CS) er proteinrammeverket som tillater ende-til-ende-kryss mellom homologe kromosomer. Det forekommer bare under pachytene av meiose I, og er det fysiske grunnlaget for kromosomal parring. Med andre ord er det det som gjør at kromosomer kan synapse og rekombinere..
Det synaptonemiske komplekset er svært bevart blant eukaryoter som gjennomgår meiose. Derfor er den evolusjonært veldig gammel, og strukturelt og funksjonelt ekvivalent i alle levende ting..
Den består av et sentralt aksialt element og to sideelementer som gjentas som tennene på en glidelås eller lukking..
Det synaptonemiske komplekset er dannet fra spesifikke punkter på kromosomene under zygoten. Disse stedene er kollinære med de der DNA-brudd oppstår der synapser og rekombinasjon vil oppleves i pachytene..
Under pachytene har vi derfor en lukket glidelås. I denne konformasjonen er spesifikke punkter definert der DNA-bånd vil bli utvekslet på slutten av scenen..
Det meiotiske synaptonemiske komplekset inneholder mange strukturelle proteiner som også finnes under mitose. Disse inkluderer topoisomerase II, kondensiner, kohesiner, samt kohesinassosierte proteiner..
I tillegg til disse er proteiner som er spesifikke og unike for meiose også til stede, sammen med proteiner fra det rekombinasjonskomplekset..
Disse proteinene er en del av rekombinosomet. Denne strukturen grupperer alle proteiner som kreves for rekombinasjon. Tilsynelatende dannes rekombinosomet ikke på delepunktene, men rekrutteres, allerede dannet, mot dem.
Chiasms er de synlige morfologiske strukturene på kromosomer der kryssoverganger forekommer. Med andre ord, den fysiske manifestasjonen av utvekslingen av DNA-bånd mellom to homologe kromosomer. Chiasms er de særegne cytomorfologiske merkene til pachyten.
I all meiose må det forekomme minst ett chiasme per kromosom. Dette betyr at hver gamete er rekombinant. Takket være dette fenomenet kunne de første genetiske kartene basert på kobling og rekombinasjon utledes og foreslås..
På den annen side forårsaker mangel på chiasmer, og derfor crossover, forvrengninger på nivået av kromosomal segregering. Rekombinasjon under pachytene fungerer da som en kvalitetskontroll av meiotisk segregering..
Men evolusjonært sett gjennomgår ikke alle organismer rekombinasjon (for eksempel mannlige fruktfluer). I disse tilfellene fungerer andre mekanismer for kromosomal segregering som ikke er avhengig av rekombinasjon..
Ved utgang av zygoten er det synaptonemiske komplekset fullstendig dannet. Dette suppleres med generering av dobbeltbånds-DNA-pauser hvor kryssoverganger blir bekreftet..
Dobbelt DNA-brudd tvinger cellen til å reparere dem. I løpet av DNA-reparasjon rekrutterer cellen rekombinosomet. Båndutveksling brukes, og som et resultat oppnås rekombinante celler.
Når det synaptonemiske komplekset er fullformet, sies det at pachytene begynner.
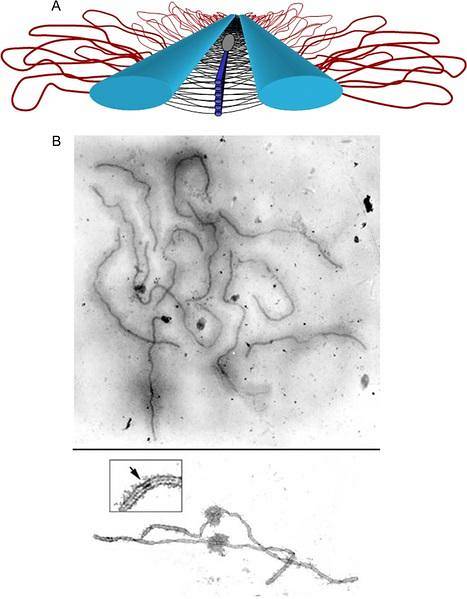
Bivalenter i synapser i pachytene samhandler i utgangspunktet gjennom det aksiale elementet i det synaptonemiske komplekset. Hver kromatid er organisert i en sløyfeorganisasjon, hvis base er det sentrale aksiale elementet i det synaptonemiske komplekset..
Det aksiale elementet til hver motpart kommer i kontakt med det andre gjennom sideelementene. Søsterkromatidaksene er svært komprimerte, og kromatinløkkene deres kommer utover fra det sentrale aksiale elementet. Sløyfeavstand (~ 20 per mikrometer) er bevart evolusjonært på tvers av alle arter.
Mot enden av pachytene, er krysninger tydelige fra noen av dobbeltbånds DNA-bruddsteder. Utseendet til delefiltrene signaliserer også begynnelsen på oppløsningen av det synaptonemiske komplekset..
Homologe kromosomer blir mer kondenserte (ser mer individuelle ut) og begynner å skilles, bortsett fra i chiasmata. Når dette skjer, slutter pachytene og diploten begynner..
Forbindelsen mellom rekombinosomet og aksene til det synaptonemiske komplekset vedvarer gjennom hele synapsen. Spesielt i rekombinogene overganger til enden av pachytenen, eller litt utover.



Ingen har kommentert denne artikkelen ennå.