De epistase, i genetikk er det studiet av samspillet mellom de forskjellige genene som koder for samme karakter. Det vil si at det er manifestasjonen av et trekk som oppstår fra interaksjonen mellom gener av gener på forskjellige steder.
Når vi snakker om relasjonene som etablerer alleler av samme gen, refererer vi til alleliske forhold. Det vil si til alleler av samme sted eller allelomorfe alleler. Dette er de velkjente interaksjonene av fullstendig dominans, ufullstendig dominans, kodominans og dødelighet mellom alleler i samme gen..
I forholdet mellom alleler på forskjellige steder, snarere tvert imot om ikke-allelomorfe alleler. Dette er de såkalte geninteraksjonene, som på en eller annen måte er epistatiske.
Epistasis gjør det mulig å analysere om uttrykket til et gen bestemmer uttrykket for et annet. I et slikt tilfelle vil et slikt gen være epistatisk på det andre; det andre ville være hypostatisk på det første. Epistasisanalyse gjør det også mulig å bestemme rekkefølgen genene som definerer den samme fenotypen virker.
Den enkleste epistasen analyserer hvordan to forskjellige gener samhandler for å gi samme fenotype. Men åpenbart kan det være mange flere gener.
For analysen av den enkle epistasen vil vi basere oss på variasjonene til proporsjonene til de klassiske dihybridkorsene. Det vil si modifikasjonene av forholdet 9: 3: 3: 1, og for seg selv.
Artikkelindeks
Denne andelen oppstår fra kombinasjonen av arveanalysen av to forskjellige tegn. Det vil si at det er produktet av kombinasjonen av to uavhengige fenotypiske segregeringer (3: 1) X (3: 1).
Når Mendel analyserte for eksempel plantevokst eller frøfarge, separerte hvert tegn 3 til 1. Da han analyserte dem sammen, selv om de var to forskjellige tegn, hver segregerte 3 til 1. Det vil si at de ble distribuert uavhengig.
Men da Mendel analyserte tegnene i par, resulterte de i de velkjente fenotypiske klassene 9, 3, 3 og 1. Men disse klassene var summer av to tegn. annerledes. Og aldri, ingen av karakterene påvirket hvordan den andre manifesterte seg.
Den forrige var forklaringen på den klassiske Mendel-andelen. Derfor er det ikke et tilfelle av epistase. Epistasis studerer tilfellene av arv av samme karakter bestemt av flere gener.
Den forrige saken, eller Mendels andre lov, var arv av to forskjellige karakterer. De senere forklarte er sanne epistatiske proporsjoner og involverer bare ikke-allelomorfe alleler..
Dette tilfellet blir funnet når den samme karakteren presenterer fire forskjellige fenotypiske manifestasjoner i et forhold på 9: 3: 3: 1. Derfor kan det ikke være en allel (monogen) interaksjon som den som fører til utseendet til fire forskjellige blodgrupper i ABO-systemet..
La oss ta et eksempel på krysset mellom et heterozygot individ med type A-blod og et heterozygot individ med type B-blod. JegTILJeg X JegBJeg. Dette vil gi oss et forhold på 1: 1: 1: 1 av individer JegTILJeg (Type A), JegTILJegB (Type AB), JegBJeg (Type B) e ii (Type O).
Tvert imot observeres et sant dominerende dobbeltepistatisk forhold (9: 3: 3: 1) i form av hanekammen. Det er fire fenotypiske klasser, men i forholdet 9: 3: 3: 1.
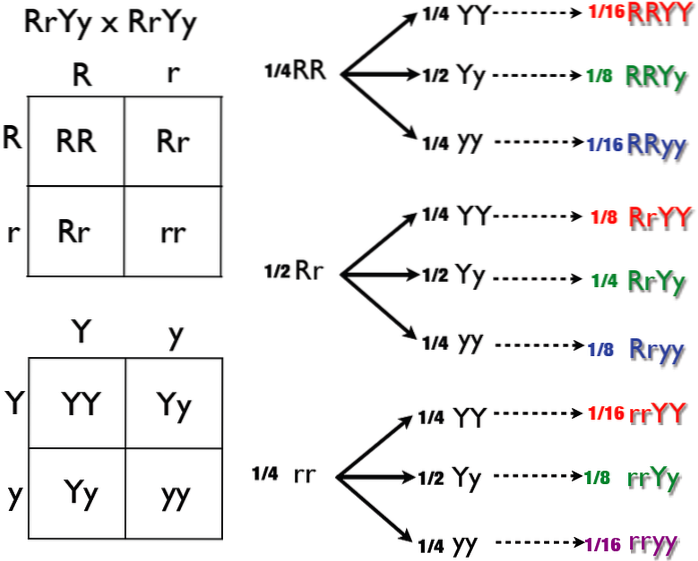
To gener deltar i bestemmelsen og manifestasjonen, la oss kalle dem R Y P. Uansett alleler R Y P vise fullstendig dominans over alleler r Y s, henholdsvis.
Av krysset RrPp X RrPp vi kan få de fenotypiske klassene 9 R_P_, 3 R_pp, 3 rrP_ og 1 rrpp. Symbolet "_" betyr at allelet kan være dominerende eller recessivt. Den tilhørende fenotypen forblir den samme.
Klasse 9 R_P_ er representert av haner med valnøttkamre, 3 R_pp av haner med rosekam. Ertekammede haner vil være klasse 3 rrP_; de av rrpp-klassen har en enkel topp.

I dobbelt dominerende epistase, oppstår hver klasse 3 fra dominanseffekten av R- eller P.-genet. Klasse 9 er representert av den hvor både R- og P-dominerende alleler manifesterer seg. Til slutt, i klasse 1 rrpp, er allelene fraværende. av begge gener.
I denne epistatiske interaksjonen undertrykker ikke ett gen manifestasjonen av et annet. Tvert imot, begge gener koder for manifestasjonen av samme trekk, men uten additiv effekt..
Derfor tillater tilstedeværelsen av minst en dominerende allel av en av de to gener fra forskjellige lokus manifestasjon av egenskapen i klasse 15. Fraværet av dominerende alleler (den dobbel recessive klassen) bestemmer fenotypen til klasse 1.
Produktene fra genene deltar i manifestasjonen av hvetekornets farge TIL meg B. Det vil si at et av disse produktene (eller begge deler) kan føre til den biokjemiske reaksjonen som forvandler forløperen til et pigment..
Den eneste klassen som ikke produserer noen av dem, er klasse 1 aabb. Derfor vil klasse 9 A_B_, 3 A_bb og 3 aaB_ produsere pigmenterte korn, og det resterende mindretallet vil ikke.
Her finner vi et tilfelle av dominant sletting av et gen (hypostatisk) på grunn av tilstedeværelsen av minst en dominerende allel av den andre (epistatisk). Formelt sett undertrykker det ene genet handlingen til det andre..
Hvis det er den dominerende undertrykkelsen av D over K, ville vi ha den samme fenotypen assosiert med klassene 9 D_K_, 3 D_kk og 1 ddkk. Klasse 3 ddK_ ville være den eneste som viser den ikke-undertrykte egenskapen.
Den dobbel recessive klassen er lagt til klasse 9 D_K_ og 3 D_kk fordi den ikke produserer det det hypostatiske genet K koder for. Ikke fordi det undertrykkes av D, som i alle fall ikke er der heller, men fordi det ikke produserer K.
Dette forholdet kalles noen ganger også dominerende og recessiv epistase. Den dominerende er den av K på D / d. Den recessive epistasen vil være den av dd på K / k.
Primrose blomster skylder for eksempel fargen sin til manifestasjonen av to gener. Gen K som koder for produksjonen av pigmentet malvidin, og genet D som koder for sletting av malvidin.
Bare planter ddKK eller ddKk (dvs. klasse 3 ddK_) vil produsere malvidin og være blå i fargen. Enhver annen genotype vil gi opphav til planter med turkise blomster.
I dette tilfellet kreves tilstedeværelsen av minst en dominerende allel av hvert gen i paret for at karakteren skal manifestere seg. La oss si at det er gener C Y P. Det vil si den homozygote recessive tilstanden til et av parets gener (DC eller pp) gjør manifestasjonen av karakteren umulig.
Med andre ord, bare klasse 9 C_P_ har minst en dominerende allel C og en dominerende allel P. For at egenskapen skal manifesteres, må de funksjonelle produktene til de to genene være tilstede.
Denne interaksjonen er epistatisk fordi mangelen på uttrykk for et gen hindrer det andre genet i å manifestere seg. Det er dobbelt, fordi det omvendte også er sant.
Et klassisk eksempel som illustrerer denne saken er erteblomstene. Plantene CCpp og plantene ccPP de har hvite blomster. CcPp-hybrider av kryssene mellom dem presenterer lilla blomster.
Hvis to av disse dihybridplanter krysses, vil vi oppnå klasse 9 C-P_, som vil ha lilla blomster. Klasse 3 C_pp, 3 ccP_ og ccpp vil være hvite blomster.

Fra andelen som er foreslått i Mendels andre lov, har vi andre tilfeller som fortjener å bli nevnt..
Vi kaller den modifiserte 9: 4: 3-forholdet recessiv epistase med god grunn. Når et gen er homozygot for det recessive genet, unngår det uttrykk for det andre genet - selv om det er dominerende.
Ta for eksempel den recessive epistasen av genotypen aa om de av genet B. Klasse 9 er den allerede anerkjente 9 A_B_. For klasse 4, til klasse 1 aabb, må de av klasse 3 aaB_ legges til, med samme fenotype. Klasse 3 vil være de i klasse 3 A_bb.
I den epistatiske interaksjonen av dupliserte gener er det observerte fenotypiske forholdet 9: 6: 1. Alle individer i klasse 9 A_B_ har minst en allel av hvert gen TIL eller B. De har alle samme fenotype.
Tvert imot, i klasse 3 A_bb og 3 aaBb er det bare dominerende alleler av enten A eller B. I dette tilfellet er det også en enkelt og samme fenotype - men forskjellig fra de andre. Til slutt er det i klasse 1 aabb ingen dominant allel av noen av genene til stede og representerer en annen fenotype.
Den kanskje mest forvirrende klassen er den dominerende epistasen, som viser det fenotypiske forholdet 12: 3: 1. Her får dominansen av A (epistatisk) over B (hypostatisk) klasse 9 A-B_ til å bli med i klasse 3 A_bb.
Fenotypen til B vil bare manifestere seg når A ikke er tilstede i klasse 3 aaB_. Den dobbeltrecessive klasse 1 aabb vil verken manifestere eller fenotypen assosiert med genet A / a ei heller med genet B / b.
Andre epistatiske fenotypiske forhold som ikke har et spesielt navn er 7: 6: 3, 3: 6: 3: 4 og 11: 5.


Ingen har kommentert denne artikkelen ennå.