De acetylkolinesterase (acetylkolinacetylhydrolase, EC 3.1.1.7) er et enzym som hovedsakelig finnes i sentralnervesystemet. Dens oppgave, som navnet antyder, er den hydrolytiske behandlingen av nevrotransmitteren acetylkolin..
Det er et enzym assosiert med cellemembranen som fungerer sammen med acetylkolinreseptoren for å formidle eksitasjonen av postsynaptiske celler, og hvis katalytiske mekanisme er overraskende rask..
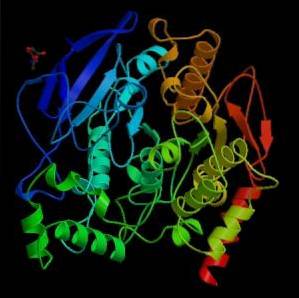
Fra et mekanisk synspunkt kan dette enzymet sees på som en serinhydrolase, og i det katalytiske domenet til det aktive stedet inneholder det triaden av aminosyrer som er karakteristiske for serinproteaser: serin, histidin og en sur rest. Den sure resten er imidlertid et glutamat, mens serinproteasene normalt har et aspartat..
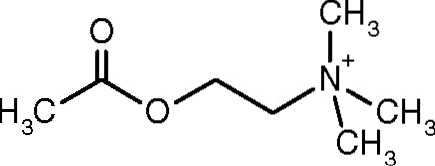
En av de første observasjonene som koblet den katalytiske aktiviteten til acetylkolinesterase med kolinergt nervevev og muskelvev ble gjort av Dale i 1914; senere er det bestemt at det også er tilstede i ikke-kolinerge nevroner og i hematopoietiske, osteogene og neoplastiske celler.
Takket være studien av forskjellige organismer er det for tiden kjent at enzymet er tilstede i membranen til forskjellige typer celler som erytrocytter, nerve- og muskelceller, elektriske organer og andre..
Artikkelindeks
Under naturlige forhold eller "in vivo ", acetylkolinesterase er et polymorft enzym som består av flere katalytiske underenheter på mer eller mindre 80 kDa, som samles for å danne en oligomer struktur (av flere underenheter).
Mengden og kompleksiteten til disse underenhetene avhenger av celletypen og arten som blir vurdert..
Noen av de mer komplekse enzymformene har katalytiske underenheter med kuleformede (G) eller asymmetriske (A) former bundet av disulfidbroer. Disulfidbroer er kovalente bindinger dannet mellom to svovelmolekyler av tiolgruppene (-SH) av to rester av aminosyren cystein.
Hver G-underenhet inneholder et enkelt aktivt sted, mens A-underenhetene generelt er preget av å ha tre strukturelle domener, nemlig: katalytiske underenheter, kollagenlignende haler rike på glycin, hydroksyprolin og hydroksylysinrester og andre. Ikke-kollagen lim (annet enn kollagen).
De asymmetriske formene av acetylkolinesterase er kjent som A12, A8 og A4, som har henholdsvis 12, 8 og 4 katalytiske underenheter..
Generelt er restene av det katalytiske domenet i det aktive stedet funnet i en "dyp" region av underenhetene, noe som kan sees på som motstridende med hensyn til den hurtige reaksjonshastigheten som katalyserer dette enzymet og den tilsynelatende utilgjengeligheten av substratet. til disse nettstedene..
Uavhengig av polymorfismen av enzymet, har både kuleformede og asymmetriske underenheter lignende katalytiske aktiviteter..
Noen andre celler enn nerveceller, som erytrocytter, produserer overveiende kuleformede, dimere acetylkolinesteraseenzymer, hovedsakelig forbundet med plasmamembranens ytre overflate..
Selv om erytrocyttenzymet har mindre strukturell kompleksitet, er det også et amfipatisk enzym, hvis aktive katalytiske domene er funnet i den store hydrofile regionen, mens det hydrofobe domenet, som inneholder det karboksylterminale området, er ansvarlig for å opprettholde det i membranen.
Mye av dagens kunnskap om sekvensen av acetylkolinesterase kom frem fra studien av enzymet Californica torpedo, en rokkefisk som lever i Stillehavet og tradisjonelt har vært brukt som modellorganisme for studier av forskjellige proteiner i nervesystemet.
Acetylkolinesterase-underenheter syntetiseres som pro-proteiner som deretter behandles for å gi opphav til modne underenheter. Hver underenhet består av et polypeptid på ca. 575 aminosyrer og 65 kDa i molekylvekt, som økes ved tilsetning av 7-8% karbohydratrester (glykosylering)..
Den katalytiske aktiviteten til det aktive stedet for underenhetene bestemmes av en serinrest i posisjon 200, som finnes i et "dypt" område av de katalytiske underenhetene..
Ulike varianter eller isoformer av enzymet finnes i organismer takket være forskjellige steder foralternativ skjøting " av RNA før pre-messenger i begge ender (5 'og 3'). Den karboksylterminale sekvensen til isoformen til hver underenhet er det som bestemmer samlingen av oligomerer med hverandre..
Acetylkolinesterase er et enzym med flere biologiske funksjoner som ikke nødvendigvis er relatert til hverandre. Fakta bekreftet av dens differensielle uttrykk under embryogenese, embryonal nevrale forlengelse, muskelutvikling og synaptogenese.
Som fremhevet ovenfor har den en viktig rolle i rask hydrolyse av acetylkolin og derfor i reguleringen av effekten i det neuromuskulære synaptiske rommet eller i de kolinerge synaptiske rommene i sentralnervesystemet..
Et eksempel på funksjonene er sammentrekning av skjelettmuskulatur, som oppstår takket være en type kjemisk synaps kjent som en motorplate, som ligger mellom et motorisk neuron og en muskelfiber..
Hundrevis av acetylkolinbelastede vesikler oppnås ved denne synapsen, som frigjøres fra motorneuronet for forplantning av en elektrisk impuls..
Denne nevrotransmisjonsprosessen er ganske komplisert, men deltakelsen av acetylkolinesterase er avgjørende for avslutningen av den synaptiske overføringen som avhenger av nevrotransmitteren acetylkolin, siden den må nedbrytes og deretter må diffundere utenfor den synaptiske kløften for å kulminere med membraneksitasjonen.
Dermed er enzymet acetylkolinesterase ansvarlig for å regulere konsentrasjonen av denne senderen i nevromotorisk synaps..
Andre "ikke-klassiske" funksjoner av enzymet er relatert til neuritogenese eller vekst av nerveceller; med prosesser for celleheft, synaptogenese, aktivering av nevroner-dopamin i substantia nigra i mellomhjernen, hematopoietiske prosesser og poietisk trombe, blant andre.
Acetylkolinesterasehemmere virker ved å forhindre at den hydrolyserer acetylkolin, og øker dermed nivået og virkningen av denne nevrotransmitteren. De kan klassifiseres i henhold til deres virkningsmekanisme som reversible og irreversible.
De er de som irreversibelt hemmer den hydrolytiske aktiviteten til acetylkolinesterase ved sin kovalente binding til serinresten i enzymets aktive sted. Denne gruppen består hovedsakelig av organofosfater.
Generelt er dette aktive forbindelser som finnes i mange insektmidler og er ansvarlige for et stort antall utilsiktet forgiftningsdødsfall. De er estere eller tioler avledet fra fosforsyre, fosfonsyre, fosfinsyre eller fosforamidinsyre.
Sarin, tabun, soman og cyclosarin er blant de mest giftige forbindelsene syntetisert av mennesker siden de kan drepe et menneske ved å indusere respiratorisk og sirkulasjonssvikt ved å blokkere acetylkolinesterase i det perifere nervesystemet..
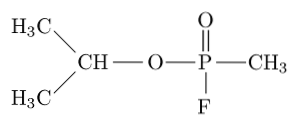
Sarin, for eksempel, er en ”nervegass” som har blitt brukt som et kjemisk våpen for terroristbruk..
Denne rekkefølgen av klassifisering grupperer konkurransedyktige og ikke-konkurransedyktige inhibitorer som virker ved forbigående og reversibel karbamylering av en serinrest i det aktive stedet, og mange er blitt syntetisert og renset fra plante- eller soppkilder..
Karbamater som fysostigmin og neostigmin er reversible hemmere som brukes som medikamenter for behandling av sykdommer som henholdsvis glaukom og myasthenia gravis..
Andre terapeutiske midler i denne gruppen brukes også til behandling av Alzheimers sykdom, Parkinsons sykdom, postoperative tarmhindringer (postoperativ ileus), blæredistens og som motgift mot antikolinerg overdose..
En interessant naturlig mekanisme mot noen acetylkolinesterasehemmende stoffer har å gjøre med deltakelse av et mindre spesifikt enzym kjent som butyrylkolinesterase.
Dette enzymet er også i stand til å hydrolysere acetylkolin, og samtidig kan det fungere som et molekylært lokkeflukt som reagerer med disse toksinene før de utøver sin negative effekt på acetylkolinesterase..
Acetylkolinesterase har vist seg å danne et stabilt kompleks med komponentene i senile plakk som er karakteristiske for patologien. Videre har noen endrede glykosyleringsmønstre av dette enzymet vært relatert til tilstedeværelsen og dannelsen av amyloide plakk i hjernen..
Mange av de reversible acetylkolinesterasehemmere har derfor blitt brukt som førstegenerasjons medisiner for behandling av denne sykdommen og andre relaterte nevrodegenerative tilstander. Disse inkluderer donepezil, rivastigmin og galantamin..


Ingen har kommentert denne artikkelen ennå.