De adenin Det er en nitrogenholdig base av purintypen, som finnes i ribonukleinsyre (RNA) og deoksyribonukleinsyre (DNA) fra levende organismer og virus. Noen av funksjonene til disse biopolymerene (RNA og DNA) er lagring, replikering, rekombinasjon og overføring av genetisk informasjon..
For å utgjøre nukleinsyrer danner først nitrogenatomet 9 i adenin en glykosidbinding med det primære karbon 1 (C1 ') av ribose (av RNA) eller 2'-deoksyribose (av DNA). På denne måten danner adenin nukleosidet adenosin eller adenosin.
For det andre danner hydroksylgruppen (-OH) av 5'-karbonet i sukkeret (ribose eller 2'-deoksyribose), av adenosin, en esterbinding med en fosfatgruppe..
Avhengig av antall tilstedeværende fosfatgrupper i levende celler, kan det være adenosin-5'-monofosfat (AMP), adenosin-5'-difosfat (ADP) og adenosin-5'-trifosfat (ATP). Ekvivalenter som har 2'-deoksyribose eksisterer også. For eksempel deoksyadenosin-5'-monofosfat (dAMP), etc..
Artikkelindeks
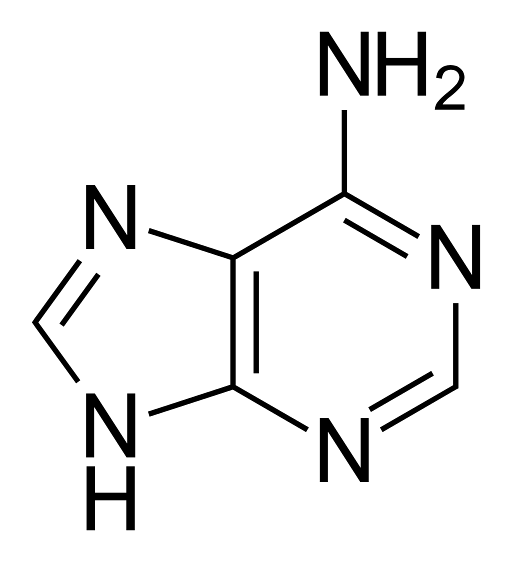
Adenin, kalt 6-aminopurin, har den empiriske formelen C5H5N5, og har en molekylvekt på 135,13 g / mol, som renses som et blekgult fast stoff, med et kokepunkt på 360 ° C..
Molekylet har en kjemisk struktur med dobbel ring med konjugerte dobbeltbindinger, som er fusjonen av et pyrimidin med en imidazolgruppe. På grunn av dette er adenin et flatt heterosyklisk molekyl..
Den har en relativ løselighet på 0,10 g / ml (ved 25 ° C), i sure og basiske vandige oppløsninger, med en pKa på 4,15 (ved 25 ° C).
Av samme grunn er den i stand til å bli detektert ved absorbans ved 263 nm (med en absorpsjonskoeffisient på E.1,2 mM = 13,2 M-1.cm-1 i 1,0 M HCl), areal av det elektromagnetiske spekteret som tilsvarer nesten ultrafiolett.
Biosyntesen av purinnukleotider er identisk i praktisk talt alle levende vesener. Det begynner med overføring av en aminogruppe fra glutamin til substratet 5-fosforibosyl-1-pyrofosfat (PRPP), og produserer 5-fosforibosylamin (PRA).
Dette er en reaksjon katalysert av glutamin-PRPP transferase, et nøkkelenzym i reguleringen av denne metabolske veien..
Etter sekvensielle tilsetninger av aminosyrene glutamin, glycin, metenyl-folat, aspartat, N10-formyl-folat til PRA, inkludert kondens og ringlukking, produseres inosin-5'-monofosfat (IMP), hvis heterosykliske enhet er hypoksantin (6-oksypurin).
Disse tilsetningene drives av hydrolyse av ATP til ADP og uorganisk fosfat (Pi). Deretter tilsettes en aminogruppe fra aspartat til IMP, i en reaksjon kombinert med hydrolyse av guanosintrifosfat (GTP), for til slutt å generere AMP.
Sistnevnte utøver kontroll over denne biosyntetiske banen gjennom negativ tilbakemelding, og virker på enzymene som katalyserer dannelsen av PRA og modifiseringen av IMP.
Som med nedbrytningen av andre nukleotider, går nitrogenbasen til adenosinnukleotider gjennom en prosess som kalles "resirkulering".
Resirkulering består av overføring av en fosfatgruppe fra PRPP til adenin, og danner AMP og pyrofosfat (PPi). Det er et enkelt trinn katalysert av enzymet adenin fosforibosyltransferase.
Adenin er en del av flere viktige molekyler i oksidativ metabolisme, som er følgende:
Under oksidativ metabolisme, NAD+ den fungerer som et elektronakseptorsubstrat (hydridioner) og danner NADH. Mens FAD er en kofaktor som aksepterer elektroner og blir FADHto.
På den annen side danner adenin nikotinamidadenindinukleotidfosfat (NADP+/ NADPH), som er involvert i å redusere stoffskiftet. For eksempel er NADPH et elektrondonatorsubstrat under lipid- og deoksyribonukleotidbiosyntese..
Adenin er en del av vitaminene. For eksempel er niacin forløperen til NAD+ og NADP+ og riboflavin er forløperen til FAD.
Adenin er en del av S-adenosylmetionin (SAM), som er en metylradikal-donor (-CH3) og deltar i metylering av adenin- og cytosinrester i prokaryoter og eukaryoter.
I prokaryoter gir metylering sitt eget DNA-gjenkjenningssystem, og beskytter dermed DNA fra sine egne restriktive enzymer..
I eukaryoter bestemmer metylering genuttrykk; det vil si at den etablerer hvilke gener som skal uttrykkes og hvilke som ikke skal. I tillegg kan adeninmetyleringer markere reparasjonssteder for skadet DNA..
Mange proteiner som binder seg til DNA, som transkripsjonsfaktorer, har aminosyrerester glutamin og asparagin som danner hydrogenbindinger med N-atomet.7 av adenin.
Adenin er en del av ATP, som er et høyenergimolekyl; det vil si at dens hydrolyse er eksergonisk, og Gibbs fri energi er en høy og negativ verdi (-7,0 Kcal / mol). I celler deltar ATP i mange reaksjoner som krever energi, for eksempel:
- Boost enzymkatalyserte endergonic kjemiske reaksjoner involvert i mellommetabolisme og anabolisme, gjennom dannelse av høyenergi-mellomprodukter eller koblede reaksjoner.
- Fremme proteinbiosyntese i ribosomer, ved å la forestring av aminosyrer med deres tilsvarende overførings-RNA (tRNA), til å danne aminoacyl-tRNA.
- Øk bevegelsen av kjemikalier over cellemembraner. Det er fire typer transportørproteiner: P, F, V og ABC. P-, F- og V-typene bærer ioner, og ABC-typen bærer underlag. For eksempel Na ATPase+/ K+, klasse P, trenger du en ATP for å pumpe inn i cellen to K+ og ut tre Na+.
- Øk muskelsammentrekning. Gir energien som leder aktinfilament glir over myosin.
- Øk kjernefysisk transport. Når beta-underenheten til den heterodimere reseptoren binder seg til ATP, interagerer den med komponenter i kjerneporekomplekset.
Adenosin fungerer som en ligand for reseptorproteiner som er tilstede i nevroner og celler i tarmepitelet, der det fungerer som et ekstracellulært eller nevromodulatorisk budskap, når endringer oppstår i cellulær energimetabolisme.
Adenin er til stede i kraftige antivirale midler som arabinosiladenin (araA), som produseres av noen mikroorganismer. I tillegg er det til stede i puromycin, et antibiotikum som hemmer proteinbiosyntese og produseres av mikroorganismer i slekten Streptomyces.
I AMP fungerer det som et substrat for reaksjoner som genererer den andre messenger sykliske AMP (cAMP). Denne forbindelsen, produsert av enzymet adenylat cyklase, er essensiell i en stor del av de intracellulære signalkaskadene, nødvendig for celleproliferasjon og overlevelse, samt betennelse og celledød..
Sulfat i fri tilstand er ikke reaktivt. Når den kommer inn i cellen blir den adenosin-5'-fosfosulfat (APS), og deretter 3'-fosfoadenosin-5'-fosfosulfat (PAPS). Hos pattedyr er PAPS donor av sulfatgrupper og danner organiske sulfatestere som de av heparin og kondroitin..
I cysteinbiosyntese tjener S-adenosylmetionin (SAM) som en forløper for syntesen av S-adenosylhomocystein, som transformeres av flere trinn, katalysert av enzymer, til cystein..
Eksperimentelt har det vist seg at oppbevaring av hydrogencyanid (HCN) og ammoniakk (NH3), under laboratorieforhold som ligner de som hersket på tidlig jord, produseres adenin i den resulterende blandingen. Dette skjer uten behov for levende celler eller cellulært materiale..
Prebiotiske forhold inkluderer fravær av fritt molekylært oksygen, en sterkt reduserende atmosfære, intens ultrafiolett stråling, store elektriske buer som de som genereres i stormer og høye temperaturer. Dette antar at adenin var den viktigste og mest utbredte nitrogenbasen dannet under prebiotisk kjemi..
Dermed vil syntesen av adenin utgjøre et sentralt trinn som vil gjøre opprinnelsen til de første cellene mulig. Disse måtte ha en membran som dannet et lukket rom, der molekylene som var nødvendige for å bygge de første biologiske polymerene som var nødvendige for selvforevaltning, ble funnet..
Adenin er sammen med andre organiske og uorganiske kjemiske forbindelser en viktig ingrediens i oppskriften som brukes i alle biokjemi-, genetikk-, molekylærbiologi- og mikrobiologilaboratorier i verden for å dyrke levedyktige celler over tid..
Dette er fordi ville normale cellevarianter kan oppdage og fange opp tilgjengelig adenin fra omgivelsene og bruke det til å syntetisere sine egne adeninnukleosider..
Dette er en form for celleoverlevelse, som sparer interne ressurser ved å syntetisere mer komplekse biologiske molekyler fra enkle forløpere tatt utenfra..
I eksperimentelle modeller av kronisk nyresykdom har mus en mutasjon i adeninfosforibosyltransferasegenet som produserer et inaktivt enzym. Disse musene administreres kommersielle løsninger som inneholder adenin, natriumcitrat og glukose, intravenøst, for å fremme deres raske utvinning..
Denne behandlingen er basert på det faktum at PRPP, den første metabolitten for purinbiosyntese, syntetiseres fra ribose-5-fosfat gjennom pentosefosfatveien, hvis utgangsmetabolitt er glukose-6-fosfat. Imidlertid er mange av disse løsningene ikke godkjent av internasjonale reguleringsorganer for menneskelig bruk..



Ingen har kommentert denne artikkelen ennå.