
De Golgi-apparatet, Også kjent som Golgi-kompleks, det er en membrancelleorganell dannet av et sett med flate vesikler stablet sammen; Disse sekkene har væske inni. Finnes i et bredt utvalg av eukaryoter, inkludert dyr, planter og sopp.
Denne organellen er ansvarlig for prosessering, emballering, klassifisering, distribusjon og modifisering av proteiner. I tillegg har den også en rolle i syntesen av lipider og karbohydrater. På den annen side forekommer syntesen av komponentene i celleveggen i planter i Golgi-apparatet..
Golgi-apparatet ble oppdaget i 1888, mens nerveceller ble studert; oppdageren, Camillo Golgi, vant Nobelprisen. Strukturen kan påvises ved sølvkromatfarging.
Først var organets eksistens tvilsom for datidens forskere, og de tilskrev Golgi-observasjonene til enkle gjenstandsprodukter av teknikkene som ble brukt.
Artikkelindeks
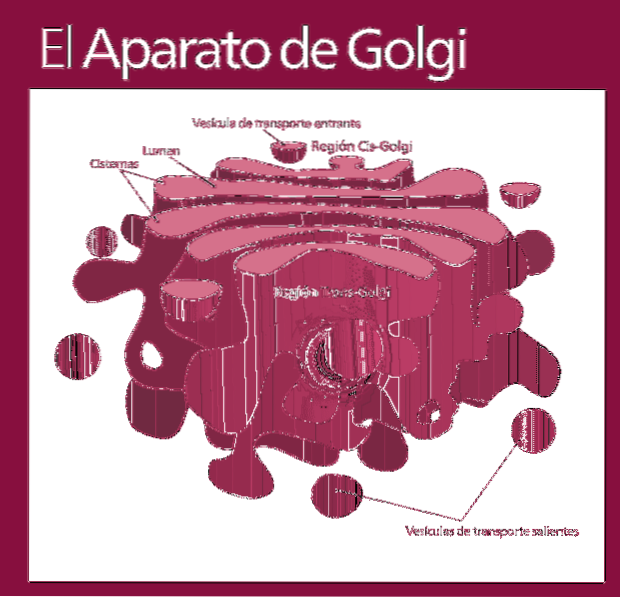
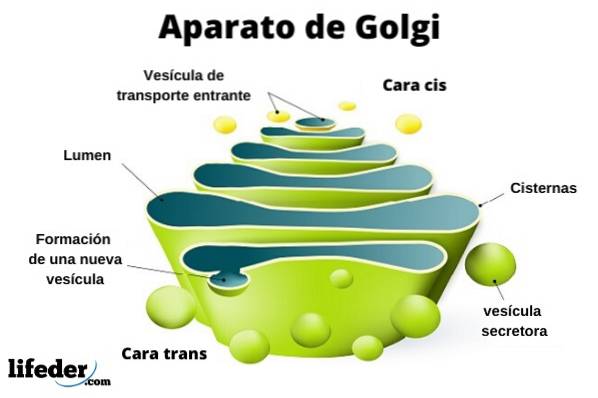
Golgi-apparatet er en eukaryot organell av en membranøs natur. Det ligner poser i stabler, selv om organisasjonen kan variere avhengig av celletype og organisme. Det er ansvarlig for modifisering av proteiner etter oversettelse.
For eksempel kan noe karbohydrat tilsettes for å danne et glykoprotein. Dette produktet er pakket og distribuert til celleområdet der det er nødvendig, slik som membran, lysosomer eller vakuoler; den kan også sendes utenfor cellen. Det deltar også i syntesen av biomolekyler.
Cytoskjelettet (spesifikt aktin) bestemmer dets plassering, og generelt ligger komplekset i et område av det indre av cellen nær kjernen og sentrosomet.
Golgi-komplekset består av et sett med flate, fenestrerte, skiveformede sekker, kalt Golgian-sisterner, med variabel tykkelse.
Disse sekkene er stablet, i grupper på fire eller seks cisterner. I en pattedyrcelle kan du finne mellom 40 og 100 batterier koblet til hverandre.
Golgi-komplekset presenterer en interessant egenskap: det er polaritet når det gjelder struktur og funksjon.
Du kan skille mellom cis ansiktet og trans ansiktet. Den første er relatert til inngangen til proteiner og ligger i nærheten av endoplasmatisk retikulum. Det andre er utgangsflaten eller produktsekresjonen De er dannet av en eller to tanker som har rørform.
Sammen med denne strukturen er vesikler som utgjør transportsystemet. Haugene med sekker er festet til hverandre i en struktur som minner om en bue eller dato.
Hos pattedyr er Golgi-komplekset fragmentert i flere vesikler under celledelingsprosesser. Blærene passerer til dattercellene og tar igjen den tradisjonelle formen av komplekset.
Organiseringen av komplekset er ikke vanlig i alle grupper av organismer. I noen celletyper er komplekset ikke strukturert som sett med cisterner stablet i grupper; tvert imot, de er lokalisert hver for seg. Et eksempel på denne organisasjonen er soppen Saccharomyces cerevisiae.
I noen encellede organismer, for eksempel toksoplasma eller trypanosom, tilstedeværelsen av bare en membranbunke er rapportert.
Alle disse unntakene indikerer at stabling av konstruksjonene ikke er viktig for å oppfylle deres funksjon, selv om nærheten mellom posene gjør transportprosessen mye mer effektiv..
På samme måte mangler noen basale eukaryoter disse cisternene; for eksempel sopp. Dette beviset støtter teorien om at apparatet dukket opp i en senere avstamning enn de første eukaryoter..
Funksjonelt er Golgi-komplekset delt inn i følgende rom: cis-nettverket, de stablede sekkene - som igjen er delt inn i midten og trans-underrommet - og trans.
Molekylene som vil bli modifisert, kommer inn i Golgi-komplekset i samme rekkefølge (cis-nettverk, etterfulgt av underkomponenter som endelig blir utskilt i trans-nettverket).
De fleste reaksjoner forekommer i de mest aktive sonene: trans- og midtre underrom.
Golgi-komplekset har som hovedfunksjon post-translationell modifisering av proteiner takket være enzymene de har inni.
Disse modifikasjonene inkluderer prosesser med glykosylering (tilsetning av karbohydrater), fosforylering (tilsetning av en fosfatgruppe), sulfatering (tilsetning av en fosfatgruppe) og proteolyse (nedbrytning av proteiner).
Videre er Golgi-komplekset involvert i syntesen av spesifikke biomolekyler. Hver av funksjonene er beskrevet i detalj nedenfor:
I Golgi-apparatet skjer modifiseringen av et protein til et glykoprotein. Den typiske sure pH i organellens indre er avgjørende for at denne prosessen skal skje normalt..
Det er en konstant utveksling av materialer mellom Golgi-apparatet med det endoplasmatiske retikulumet og lysosomene. I det endoplasmatiske retikulumet gjennomgår proteiner også modifikasjoner; disse inkluderer tilsetning av et oligosakkarid.
Når disse molekylene (N-oligosakkarider) kommer inn i Golgi-komplekset, mottar de en rekke ytterligere modifikasjoner. Hvis skjebnen til dette molekylet skal bæres utenfor cellen eller mottas i plasmamembranen, oppstår spesielle modifikasjoner.
Disse modifikasjonene inkluderer følgende trinn: fjerning av tre mannoserester, tilsetning av N-acetylglukosamin, fjerning av to mannoser, og tilsetning av fukose, ytterligere to N-acetylglukosamin-, tre galaktose- og tre sialinsyrerester..
I kontrast til dette blir proteinene som er bestemt for lysosomer modifisert på følgende måte: det er ingen fjerning av mannoser som et første trinn; i stedet oppstår fosforylering av disse restene. Dette trinnet skjer i cis-regionen av komplekset.
Deretter fjernes N-acetylglukosamin-gruppene og etterlater mannosene med fosfatet tilsatt til oligosakkaridet. Disse fosfater indikerer at proteinet må være spesielt rettet mot lysosomer..
Reseptorene som er ansvarlige for å gjenkjenne fosfatene som indikerer deres intracellulære skjebne, ligger i trans-nettverket.
I Golgi-komplekset oppstår syntesen av glykolipider og sfingomyelin ved bruk av ceramid (tidligere syntetisert i endoplasmatisk retikulum) som opprinnelsesmolekyl. Denne prosessen er i strid med den for resten av fosfolipidene som utgjør plasmamembranen, som er avledet fra glyserol..
Sphingomyelin er en klasse av sfingolipid. Det er en rikelig komponent av pattedyrsmembraner, spesielt nerveceller, der de er en del av myelinskeden.
Etter syntesen blir de transportert til deres endelige plassering: plasmamembranen. Polarhodene deres ligger mot utsiden av celleoverflaten; disse elementene har en spesifikk rolle i cellegjenkjenningsprosesser.
I planteceller bidrar Golgi-apparatet til syntesen av polysakkaridene som utgjør celleveggen, spesielt hemicellulose og pektiner. Gjennom vesikulær transport blir disse polymerene ført utenfor cellen.
I grønnsaker er dette trinnet avgjørende, og omtrent 80% av aktiviteten til retikulum er tilordnet syntesen av polysakkarider. Faktisk har hundrevis av disse organellene blitt rapportert i planteceller..
De forskjellige biomolekylene - proteiner, karbohydrater og lipider - overføres til deres cellulære destinasjoner av Golgi-komplekset. Proteiner har en slags "kode" som er ansvarlig for å informere destinasjonen den tilhører.
De transporteres i vesikler som forlater transnettverket og reiser til det spesifikke celleområdet.
Proteiner kan føres til membranen på en spesifikk konstituerende vei. Det er derfor det er en kontinuerlig inkorporering av proteiner og lipider i plasmamembranen. Proteiner hvis endelige destinasjon er Golgi-komplekset beholdes av dette.
I tillegg til den konstituerende banen, er andre proteiner bestemt for det ytre av cellen og skjer gjennom signaler fra omgivelsene, det være seg hormoner, enzymer eller nevrotransmittere..
For eksempel, i bukspyttkjertelceller, er fordøyelsesenzymer pakket i vesikler som bare utskilles når nærvær av mat oppdages..
Nyere forskning rapporterer om eksistensen av alternative veier for membranproteiner som ikke passerer gjennom Golgi-apparatet. Imidlertid disse stiene omgå "Ukonvensjonelt" debatteres i litteraturen.
Det er fem modeller som forklarer handel med proteiner i apparatet. Den første involverer trafikk av materiale mellom stabile rom, hver har de nødvendige enzymene for å oppfylle spesifikke funksjoner. Den andre modellen innebærer modning av sisternene gradvis.
Den tredje foreslår også modning av posene, men med innarbeidelsen av en ny komponent: den rørformede transporten. I følge modellen er tubuli viktig i trafikken i begge retninger.
Den fjerde modellen foreslår at komplekset fungerer som en enhet. Den femte og siste modellen er den nyeste og fastholder at komplekset er delt inn i forskjellige rom.
I visse celletyper har Golgi-komplekset spesifikke funksjoner. Cellene i bukspyttkjertelen har spesialiserte strukturer for utskillelse av insulin.
De forskjellige blodtypene hos mennesker er et eksempel på differensielle glykosyleringsmønstre. Dette fenomenet forklares med tilstedeværelsen av forskjellige alleler som koder for glukotransferase.



Ingen har kommentert denne artikkelen ennå.