Calmodulin er et begrep som kommer til å bety "kalsiummodulert protein", og refererer til et lite intracellulært protein som har egenskapen til å binde seg til kalsiumionen (Ca ++) og å formidle mange av dets intracellulære handlinger. Opprinnelsen til ordet kommer fra kombinasjonen av de engelske ordene 'kalsium', 'modulert' og 'protein' som når de settes sammen, stammer fra KALMcium MODULated proteI.
Blant mineralelementene som blir en del av konstitusjonen av dyreorganismer, er kalsium, etterfulgt av fosfor, den langt vanligste siden bein dannes ved avsetning i matrisen av store mengder mineralsalter dannet fra dette ionet..

Selvfølgelig er disse mineralsaltene av kalsium essensielle for konstitusjon og konformasjon av skjelettsystemet hos virveldyr, men det er den ioniserte kalsiumformen (Ca ++) i oppløsning i kroppsvæsker som får en relevant fysiologisk betydning for livet til dyr. organismer.
Denne kationen, med to overskytende positive elektriske ladninger i strukturen, kan fungere som en strømbærer ved å bevege seg gjennom cellemembranen og endre nivået av elektrisk potensial i mange av de eksiterende cellene i kroppen, hovedsakelig i hjertemuskelen..
Men av større fysiologisk relevans er det faktum at mange cellulære regulatoriske reaksjoner utløst av eksterne stimuli, slik som nevrotransmittere, hormoner eller andre fysiske eller biokjemiske faktorer, er arter av metabolske kaskader der flere proteiner sekvensielt deltar, hvorav noen er enzymer som krever kalsium for aktivering eller inaktivering.
Det sies da i disse tilfellene at kalsium fungerer som en andre budbringer i en metabolsk kaskade bestemt til å ha et endelig resultat som vil bli som den cellulære responsen som er nødvendig for å tilfredsstille et behov oppdaget på et annet nivå som er forskjellig fra selve cellen, og at det krever henne akkurat det svaret.
Kalsium kan handle direkte på sitt biokjemiske mål for å påvirke dets aktivitet, men det krever ofte deltakelse av et protein som det må binde seg for å utøve sin effekt på proteinet / proteinene som skal modifiseres. Calmodulin er et av disse mediatorproteinene.
Artikkelindeks
Calmodulin, svært allestedsnærværende, siden det uttrykkes i nesten alle celletyper av eukaryote organismer, er et lite surt protein med en molekylvekt på ca. 17 kDa, hvis struktur er sterkt konservert blant arter..
Det er et monomert protein, det vil si at det består av en enkelt polypeptidkjede, som i endene ender i form av kuleformede domener forbundet med en alfa-helix. Hvert kuledomene har to motiver kjent som EF-hånden. EF hånd) som er typiske for kalsiumbindende proteiner.
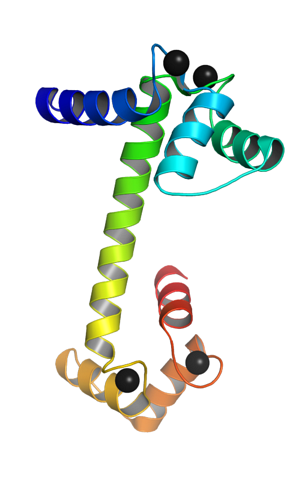
Disse "EF-hånden" topologiske motivene representerer en slags supersekundære strukturer; er knyttet til hverandre, i hvert globulært domene, av en region med stor fleksibilitet, og i hver av dem er det et bindingssted for Ca ++, som gir 4 nettsteder totalt for hvert calmodulinmolekyl.
Bindingen av positivt ladede kalsiumioner er mulig ved tilstedeværelsen av aminosyrerester med negativt ladede sidekjeder ved kalsiumbindingsstedene til calmodulin. Disse restene er tre aspartater og ett glutamat..
Alle funksjonene som hittil er kjent for calmodulin er innrammet i en gruppe handlinger som fremmes av økningen i cytosolisk kalsium produsert av dets inntreden fra det ekstracellulære rommet eller dets utgang fra de intracellulære avsetningene: mitokondrier og endoplasmatisk retikulum.
Mange av kalsiumens handlinger utføres av dette ionet ved å virke direkte på dets målproteiner, som kan ha forskjellige typer og funksjoner. Noen av disse proteinene kan ikke påvirkes direkte, men krever kalsium for å binde seg til calmodulin, og det er dette komplekset som virker på proteinet påvirket av ionet..
Disse målproteinene sies å være avhengig av kalsium-kalmodulin og inkluderer dusinvis av enzymer, så som proteinkinaser, proteinfosfataser, nukleotidsyklaser og fosfodiesteraser; alle involvert i et utall fysiologiske funksjoner, inkludert:
- Metabolisme
- Partikkeltransport
- Innflytelsesevne
- Sekresjonen av stoffer
- Befruktning av eggløsningene
- Genuttrykk
- Celleproliferasjon
- Den strukturelle integriteten til celler
- Intercellulær kommunikasjon osv.
Blant de kalmodulin-avhengige proteinkinaser er: myosin lettkjedekinase (MLCK), fosforylasekinase og Ca ++ / calmodulin-kinaser I, II og III.
Således blir informasjonen kodet av kalsiumsignaler (økning eller reduksjon i dens intracellulære konsentrasjon) "dekodet" av dette og andre kalsiumbindende proteiner, som omdanner signalene til biokjemiske endringer; Med andre ord er calmodulin et mellomprotein i kalsiumavhengige signalprosesser.
Calmodulin er et veldig allsidig protein, da dets "mål" proteiner er betydelig forskjellige i form, sekvens, størrelse og funksjon. Siden det er et protein som fungerer som en "sensor" for kalsiumioner, avhenger virkningsmekanismen av endringene som er indusert i struktur og / eller konformasjon når det binder seg til fire av disse ionene..
Dens virkningsmekanismer kan eksemplifiseres ved å kort gjennomgå dens deltakelse i et par fysiologiske prosesser som sammentrekning av glatt muskulatur i innvollene og tilpasning til lukt som lages av hårcellene i olfaktorisk slimhinne i nesen..
Skjelett- og hjertemuskulær sammentrekning utløses når økningen i cytosolisk Ca ++ når nivåer over 10-6 mol / l og dette ionet binder seg til troponin C, som gjennomgår allosteriske endringer som påvirker tropomyosin. I sin tur beveger tropomyosin og eksponerer dets myosinbindende steder i aktin, noe som resulterer i at den kontraktile prosessen blir utløst.
Troponin C eksisterer ikke i glatt muskulatur, og økningen i Ca ++ over det angitte nivået fremmer bindingen med kalmodulin. Ca-calmodulin-komplekset aktiverer myosin lettkjedekinase (MLCK), som igjen fosforylerer denne lette kjeden, aktiverer myosin og utløser kontraktil prosess..
Økningen i Ca ++ skjer gjennom inngangen fra utsiden eller utgangen fra det sarkoplasmatiske retikulumet ved virkningen av inositoltrifosfat (IP3) frigitt av fosfolipase C i kaskaden aktivert av reseptorer koblet til Gq-protein. Avspenning oppstår når Ca ++, ved hjelp av transportører, fjernes fra cytosolen og returnerer til opprinnelsesstedene.
En viktig forskjell mellom begge typer sammentrekning er at i strierte muskler (hjerte og skjelett) Ca ++ induserer allosteriske endringer ved å binde seg til proteinet, troponin, mens i glatte muskler er endringene produsert av Ca-calmodulin kovalente og involverer fosforylering av myosin.
Derfor, når handlingen av Ca ++ er avsluttet, er det nødvendig med deltakelse av et annet enzym for å fjerne fosfatet tilsatt av kinasen. Dette nye enzymet er myosin lettkjede fosfatase (MLCP) hvis aktivitet ikke avhenger av calmodulin, men er regulert av andre veier.
I virkeligheten opphører ikke den kontraktile prosessen med glatt muskulatur helt, men graden av sammentrekning forblir på et mellomnivå som et resultat av balansen mellom begge enzymers handlinger, MLCK kontrollert av Ca ++ og calmodulin, og MLCP underlagt andre regulatoriske kontroller.
Den luktende følelsen utløses når olfaktoriske reseptorer som er lokalisert i cilia av celler som ligger på overflaten av lukteslimhinnen, aktiveres..
Disse reseptorene er koblet til et heterotrimert G-protein kjent som "Golf" (olfaktorisk G-protein), som har tre underenheter: "αolf", "ß" og "γ".
Når olfaktoriske reseptorer aktiveres som respons på en lukt, dissosierer underenhetene til dette proteinet, og "aolf" -enheten aktiverer enzymet adenylcyklase og produserer syklisk adenosinmonofosfat (cAMP)..
CAMP aktiverer CNG-lignende kanaler (aktivert av sykliske nukleotider) for kalsium og natrium. Disse ionene kommer inn i cellen, depolariserer den og forårsaker initiering av handlingspotensialer hvis frekvens vil bestemme luktens intensitet..
Kalsiumet som kommer inn, som har en tendens til å depolarisere cellen, har en antagonistisk effekt av negativ tilbakemelding, noe senere, ved å binde seg til calmodulin og mellom dem å lukke kanalen og eliminere den depolariserende stimulansen, selv om den luktende stimulansen vedvarer. Dette kalles sensortilpasning.
Planter reagerer også på forskjeller i den intracellulære konsentrasjonen av kalsiumioner gjennom proteinet calmodulin. I disse organismene deler kalmoduliner mange strukturelle og funksjonelle egenskaper med sine kolleger hos dyr og gjær, selv om de er forskjellige i noen funksjonelle aspekter..
For eksempel binder calmodulin i planter til korte peptidsekvenser i målproteinene, og induserer strukturelle endringer som endrer deres aktiviteter som respons på interne variasjoner i kalsium..
I hvilken grad calmodulin kontrollerer prosesser som er analoge med de som forekommer hos dyr i planter, er det fortsatt et diskusjonsspørsmål i dag..

Ingen har kommentert denne artikkelen ennå.