
De ionekanaler de er hule membranstrukturer som danner ledninger eller porer som krysser tykkelsen på membranen og kommuniserer det ytre av cellen med dens cytosol og omvendt; noen kan ha et portanlegg som regulerer åpningen.
Disse kanalene er fylt med vann og kontrollerer passering av spesifikke ioner fra den ene siden av membranen til den andre. De dannes av noen proteiner som er typiske for cellemembraner som danner sylindriske rørformede strukturer som krysser dem i bredden..
Transportmekanismene over disse membranene kan i stor grad klassifiseres i passiv eller aktiv transport. Passive er de som tillater passering av stoffer til fordel for konsentrasjonsgradientene, mens aktive krever energiforbruk, siden de flytter stoffer mot konsentrasjonsgradientene..
Ionekanaler utgjør en passiv transportmekanisme som kan klassifiseres i henhold til deres spesifisitet, det vil si i henhold til den type ion som de lar passere gjennom, eller i henhold til måten de åpnes eller lukkes på..

Hovedfunksjonen til disse membrantransportsystemene er å tillate regulert passering av stoffer inn i eller ut av celler og dermed opprettholde intracellulære konsentrasjoner av ioner og andre stoffer..
Tilstedeværelsen av cellemembraner og ionekanaler er viktig for å opprettholde konsentrasjonsforskjeller mellom det intracellulære og ekstracellulære mediet, noe som er relevant fra mange synsvinkler..
Ionkanaler, spesielt de avhengige ligandene, er veldig viktige i farmakologi og medisin, siden mange medikamenter kan etterligne funksjonene til naturlige ligander og dermed binde seg til kanalen, åpne eller lukke den, alt etter omstendighetene..
Andre legemidler er i stand til å blokkere bindingsstedet og dermed forhindre virkningen av den naturlige liganden.
Artikkelindeks
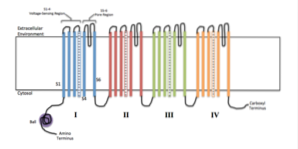
Strukturen til ionekanaler består av spesifikke transmembrane proteiner som har en rørform og etterlater en pore eller et hull som tillater kommunikasjon mellom innsiden og utsiden av cellen eller mellom intracellulære rom (organeller)..
Hver ionekanal involverer et spesifikt strukturelt membranprotein, og mer enn 100 gener som koder for spesifikke ionekanaler er beskrevet..
For natriumkanalen, for eksempel, 10 gener kalt SCN som koder for forskjellige proteiner distribuert i forskjellige vev med spesielle funksjoner og strukturer.
På samme måte er det blitt beskrevet en betydelig mengde gener som koder for forskjellige proteiner som utgjør kaliumkanaler som tilhører forskjellige familier og som har forskjellige aktiverings-, åpnings- og inaktiveringsmekanismer..
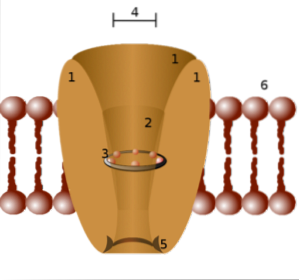
Vanligvis er en funksjonell ionekanal assosiert med en membran sammensatt av sammenstillingen av 4 til 6 lignende polypeptid-underenheter (homo-oligomerer) eller forskjellige (hetero-oligomerer) som danner en sentral pore mellom dem..
Hver underenhet varierer i henhold til karakteristikken og egenskapene til kanalen, siden mange er spesifikke for visse ioner og har forskjellige åpnings- og lukkemekanismer..
Noen kanaler består av en enkelt polypeptidkjede organisert i gjentatte motiver som går gjennom flere ganger tykkelsen på membranen og fungerer som ekvivalenten til en proteinunderenhet..
I tillegg til disse underenhetene, som i litteraturen er kjent som α-underenheter, har noen ionekanaler også en eller flere hjelpeunderenheter (ß eller γ) som regulerer åpning og lukking..
Spesifisiteten til hver kanal er relatert til diameteren på porene som dannes av transmembrane proteiner og sidekjedene (─R) til aminosyrene som utgjør dem..
På denne måten er det kanaler som bare lar natrium, kalium, kalsiumioner passere og så videre, siden sidekjedene fungerer som en "sil".
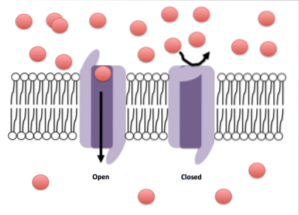
Et annet viktig trekk ved mange kanaler er portene. Kanaler med disse egenskapene kan åpne eller lukke mot lokale endringer som oppstår i membranmikromiljøet som omgir kanalen..
Avhengig av typen kanal, kan disse endringene være mekaniske, termiske (temperaturendringer), elektriske (spenningsendringer) eller kjemiske (binding av en ligand).
Imidlertid, i de såkalte passive ionkanalene, som er de som forblir åpne og tillater spesifikk passering av visse ioner, har disse strukturene ikke porter eller er følsomme for ligander eller andre typer stimuli..
I andre ionekanaler, som er følsomme for tilstedeværelse eller binding av ligander, er det et bindingssted for ligand enten på den ekstracellulære siden eller mot cellecytosolen, og i disse tilfellene har porene eller kanalene en gate som kan åpnes eller lukkes i henhold til statusen til liganden din.
I tilfelle å ha et ligandsted i den intracellulære delen, har disse kanalene vanligvis andre budbringere som ligander. Et eksempel på ionekanaler som åpnes eller lukkes av andre messenger-mekanismer er at av olfaktoriske reseptorer:
Duftmolekyler binder seg til reseptorene på den ekstracellulære siden. Disse reseptorene er i sin tur festet til et aktivert G-protein som i sin tur aktiverer proteinadenylsyklasen som danner cAMP, som er en andre messenger..
CAMP binder seg til et intracellulært bindingssted for noen kalsiumkanaler, noe som resulterer i åpning og innføring av kalsium i cellen.
Som om det var en dominoeffekt, binder kalsium seg til et ligandsted i en annen klorkanal, som genererer åpningen og utgangen av dette ionet, og forårsaker avpolarisering av luktecellen.
Det er viktig å markere at endringene generert av ligandene eller stimuli som påvirker ionekanalene tilsvarer konformasjonsendringer av proteinene som utgjør strukturen til kanalen..
Med andre ord, konformasjonsendringene som kan bevege en gate og lukke eller åpne en kanal, er ikke noe mer enn å nærme seg eller distansere protein-underenhetene som komponerer den..
Noen kanaler, spesielt spenningsavhengige kanaler, kan komme inn i en ildfast tilstand der den samme spenningsendringen som aktiverte dem nå ikke lenger aktiverer dem..
For eksempel i spenningsstyrte kalsiumkanaler åpner spenningsendringen kanalen og kalsium kommer inn, og når den er inne i cellen, binder det samme ionet til et kalsiumkanalbindingssted og kalsiumkanalen lukkes..
En annen form for reversibel inaktivering av kalsiumkanalen som forklarer dens ildfasthet etter aktivering er kanalfosforylering på grunn av økt intern kalsiumkonsentrasjon..
Det vil si at en kalsiumkanal kan inaktiveres irreversibelt på grunn av tilstedeværelsen av patologisk høye konsentrasjoner av ionet, som formidler rekruttering av spaltningsenzymer av andre kalsiumaktiverte proteiner..
Ligandregulerte kanaler kan komme inn i en ildfast tilstand når de utsettes for liganden i lang tid, og denne mekanismen kalles desensibilisering..
Narkotika, gift og giftstoffer kan påvirke reguleringen av ionekanaler, lukke eller holde dem åpne eller i noen tilfeller okkuperer ligandstedet og dermed forstyrre funksjonen..
Ionekanaler har en rekke funksjoner, direkte eller indirekte.
- De er ansvarlige for å regulere strømmen av ioner gjennom plasma og organellmembraner i alle celler..
- De tillater eksistensen av en kontroll over de intracellulære konsentrasjonene av de forskjellige ionene.
- I nevroner og muskelceller kontrollerer ionekanaler variasjonene i membranpotensial som oppstår under handlingspotensialer og under postsynaptiske potensialer for effektorceller..
- Kalsiumkanalene som genererer nettoflukt av kalsium i det intracellulære rommet er ansvarlige for aktivering av mange enzymer og proteiner som deltar i mange metabolske prosesser..
- Likeledes initierer økningen i kalsium på grunn av en økning i transporten frigjøringsmekanismen for nevrotransmittere til det synaptiske rommet til nevroner..
- Derfor er funksjonen til ionekanaler også relatert til mekanismene for cellulær kommunikasjon..
Som nevnt ovenfor kan membrantransportmekanismer være aktive eller passive, avhengig av om de bruker energi fra cellen der de befinner seg. Passive mekanismer klassifiseres i enkel diffusjon og letter diffusjon.
Enkel diffusjon tillater passering gjennom fosfolipidstrukturen i membranen av fettløselige molekyler av liten størrelse, med apolære egenskaper og uten kostnad..
For eksempel passerer gasser som oksygen (O2) og karbondioksid (CO2), etanol og urea, for å nevne noen, til fordel for deres konsentrasjonsgradient.
Tilrettelagt diffusjon er en som lettes av proteiner, og det er to typer av denne passive transportmekanismen: ionekanaler og transportproteiner eller transportproteiner..
Ionkanaler er den mekanismen som brukes mest av celler for transport av ioner som ikke kan passere ved enkel diffusjon, enten fordi de har en elektrisk ladning og fosfolipidene i membranen frastøter dem på grunn av deres størrelse og polaritet eller andre egenskaper..
Diffusjon muliggjort av transportørproteiner brukes til å transportere større stoffer med eller uten ladning, for eksempel glukose og annet sukker..
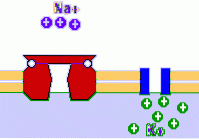
Aktiv membrantransport er den som oppstår mot konsentrasjonsgradienten til det oppløste stoffet som transporteres og krever forbruk av energi i form av ATP. Blant transportørene av denne typen er pumper og vesikulær transport.
Som et eksempel på pumpene er natrium / kaliumpumpen, som fjerner tre natrium og introduserer to kalium. Det er også kalsiumpumper.
Eksempler på vesikulær transport er endocytose, eksocytose, pinocytose og fagocytose; alle disse aktive transportmekanismene.
Fra dette punktet vil det henvises til ionekanalene som tillater passering av ioner gjennom en membran til fordel for deres konsentrasjonsgradienter, det vil si at de er passive transportkanaler..
Generelt er hver av disse kanalene spesifikke for et enkelt ion, med unntak av noen få kanaler som tillater transport av par av ioner..
En måte å klassifisere ionekanaler er ved å gruppere dem i henhold til mekanismen som er ansvarlig for åpningen. Dermed har passive kanaler, spenningsregulerte kanaler (spenningsavhengig), ligandregulerte kanaler og kanaler regulert av mekaniske stimuli blitt beskrevet..
- Passive kanaler: de er kanaler som er permanent åpne og ikke reagerer på noen form for stimulans; disse er spesifikke for visse ioner.
- Spenningsavhengige kanalerDisse kan åpne eller lukke (avhengig av kanal) i møte med endringer i membranspenningen. De er veldig viktige for cellesignalering, spesielt i sentralnervesystemet hos pattedyr.
- Ligand-avhengige kanaler: Også kalt ligand-gated eller ligand-regulerte kanaler, de er mye distribuert i forskjellige menneskelige kroppsceller, men i nervesystemet utgjør de de ionekanalene som aktiveres av nevrotransmittere og er essensielle for synaptisk overføring og intercellulær signalering.
Eksempel på ligandavhengige ionekanaler aktivert av nevrotransmittere er natrium / kaliumkanaler aktivert av glutamat..
Aktivering av kolinerge reseptorer, i dette tilfellet binding av acetylkolin til den postsynaptiske membran (kanalligand), åpner ligand-avhengige natriumkanaler og tillater innføring av dette ionet etter dets konsentrasjonsgradient..
- Kanaler regulert av mekaniske stimuli: Dette er kanaler som kan aktiveres av belastning eller trykk. Disse mekaniske kreftene overføres til kanalen gjennom cytoskelettet og kanalen åpnes.
Ingen har kommentert denne artikkelen ennå.