
De sentromerer de er grunnleggende kromosomale strukturer som holder søsterkromatider sammen under celledeling. I tillegg er dette stedet der mikrotubuli i spindelen blir sammen for å skille kromosomene på slutten av celledeling..
Sentromerer ble først beskrevet i 1882 av legen og forskeren Walther Flemming (1843-1905), da han utførte en detaljert karakterisering av celledeling..
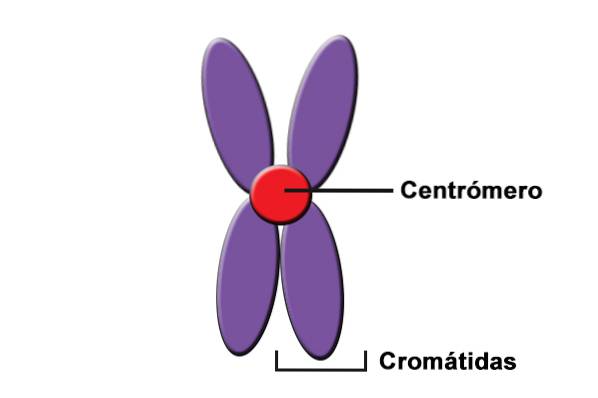
Sentromerer er også kjent som "adhesjonsregioner" eller "kinetochore". Imidlertid er det nå kjent at dette er bindingsstedene til komplekset av DNA og proteiner som danner kinetokoren..
Funksjonen til sentromeren i alle levende vesener er den samme, men hver art viser unike egenskaper, og det kan være interspesifikke forskjeller når det gjelder struktur, størrelse og kompleksitet..
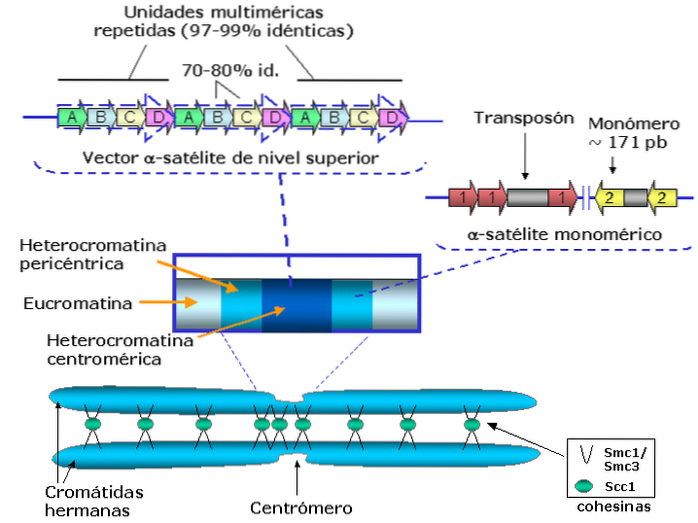
For forskere er studiet av sentromeren ikke en lett oppgave, siden "planter" eller "regioner" er i planter og dyr inneholdt i deler av det satellittlignende genomet (veldig repeterende), noe som gjør det vanskelig å kartlegge ved hjelp av teknikker for konvensjonell sekvensering.
Mutasjoner i den sentromere regionen har alvorlige fysiologiske implikasjoner hos mennesker. Avvik i strukturen og funksjonene er dødelige eller assosiert med medfødte og ervervede sykdommer, med kreft, infertilitet og fødselsforstyrrelser..
Sentromerer er deler av kromosomer som inneholder svært gjentatte regioner av DNA i form av heterokromatin. Disse regionene er spesialisert i forening og segregering av søsterkromatider under celledeling..
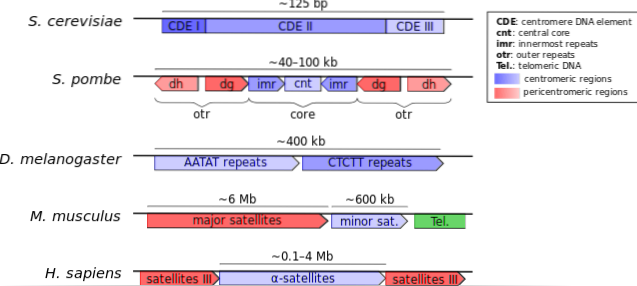
Generelt inneholder sentromerer de mest "eldste" DNA-sekvensene, ordnet suksessivt og nær grensen eller grensen mellom heterokromatin og eukromatin, det vil si at sentromerer er svært heterokromatiske regioner.
Sentromere sekvenser klassifiseres regelmessig i to typer: satellitt-DNA og transponerbare elementer. Begge typer sekvenser representerer det meste av DNA som finnes i sentromerer..
Siden sentromerer er en del av kromosomkromatin, dannes de av et kompleks av DNA og histonproteiner som favoriserer deres "emballasje".
Nukleosomene i de sentromere regionene har imidlertid ikke histon H3-proteinet; i stedet har de en variant som fagfolk har identifisert som, som er sentromerspesifikk.
Dette histonlignende proteinet varierer betydelig mellom forskjellige arter. Hos pattedyr er dette kjent som CENP-A, i leddyr kalles det CID og hos sopp og gjær har det blitt kalt Cse4.
Takket være de spesifikke forskjellene mellom CENH3-proteinet i sentromerer, blir dets egenskaper og egenskaper brukt til å identifisere arter, spesielt den sentromere regionen i kromosomene..
Plasseringen til sentromeren på kromosomene visualiseres i karyotyper som en "innsnevring" som vanligvis kalles en "primær innsnevring".
I noen organismer finnes ikke sentromerer i en region, men er ganske "diffuse", slik at fibrene i spindelen kan sammenføyes langs hele kromosomet. Disse kromosomene er kjent som diffus sentromer.
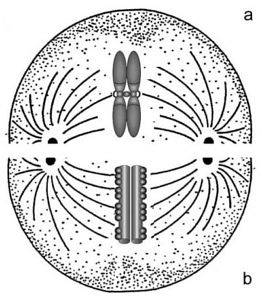
Tvert imot, hvis sentromeren er nær en av endene av et kromosom, vil denne, når den skilles fra søsterkromatiden, ha en "J" -form under segregering. På samme måte, hvis sentromeren er plassert i endene av et kromosom, vil separasjonen gi det et "stivt stang" -utseende..
Det er viktig å nevne at posisjonen til sentromeren på et kromosom indikerer forholdet mellom lengden på de to armene (den korte eller "p" og den lange eller "q"). Dette forholdet er ganske spesifikt for hver type kromosom..
I henhold til posisjonen til sentromeren gjenkjennes tre typer kromosomer:
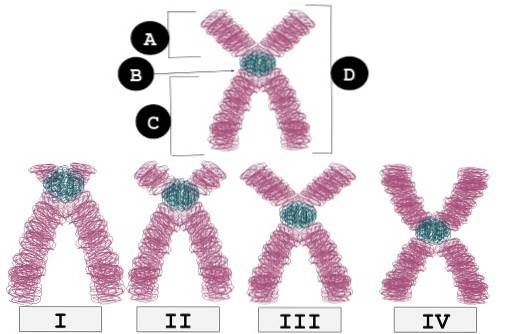
Disse kromosomene har sentromeren på slutten av en av de to kromatin “armene”. De er de som beveger seg i form av stive stenger under segregering mot polene i celledeling.
I denne typen kromosomer forskyves sentromeren mer mot den ene enden enn mot den andre. Når cellen deler seg og kromosomene skilles, er de akrosentriske kromosomene de som får en "J" -form..
Metasentriske kromosomer har sentromerer plassert gjennom hele midten av kromosomet, og skiller to like lange armer. På grunn av plasseringen av deres sentromer, skiller metasentriske kromosomer ut i V-form under anafase av celledeling.
Sentromerer er det universelle middel for effektiv utskillelse av kromosomer i alle eukaryote organismer. De er bindingsstedene for mikrotubuli for å utøve den nøyaktige mekaniske kraften for å skille kromosomer eller kromatider under meiose eller mitose..
De spesifikke funksjonene til sentromer er vedheft og separasjon av søsterkromatider, fiksering av mikrotubuli, bevegelse av kromosomer under segregering mot datterceller, etablering av heterokromatin og representerer i tillegg et kontrollpunkt for mitose.
Hos pattedyr finnes CENP-lignende proteiner i sentromeren heterokromatin. Disse kan være av tre typer CENP-A, CENP-B og CENP-C, som alle deltar i samlingen av kinetochore.
Fraværet av CENP-C-proteinet kan forårsake alvorlige feil i segregeringen av kromosomer, siden dette er et protein som har DNA-bindende og "selvassosiasjon" -egenskaper og er direkte relatert til segregering av kromosomer og kinetochore-funksjonsfeil.
Det er for tiden kjent at noen regioner av sentromerer er transkripsjonsmessig aktive. Disse koder for små interfererende RNA, som deltar i transkripsjonslyddingen av noen regioner i genomet..
Disse små dobbeltbånds-RNA-transkripsjonene fra de pericentromere regionene er essensielle for montering av heterokromatin og er transkripsjonsregionene for å regulere trinnene før celledeling..



Ingen har kommentert denne artikkelen ennå.