De erytroblaster de er forløperceller til erytrocytter av virveldyr. Reduksjonen i oksygenkonsentrasjonen i vevet vil fremme cellulære differensieringshendelser i disse cellene som vil gi opphav til modne erytrocytter. Settet med alle disse hendelsene er kjent som erytropoiesis.
Under erytropoiesis øker hemoglobinsyntese. Et rikelig protein i erytrocytter som formidler oksygentilførsel til vev og avgiftning av karbondioksid fra vev, et celletoksisk avfallsprodukt fra cellulær respirasjon.
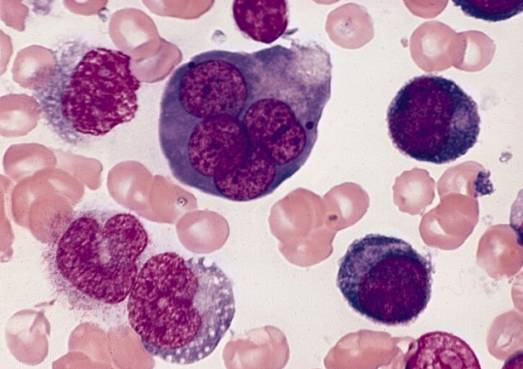
Feil i differensieringsprosessen av erytroblaster gir opphav til et sett med blodpatologier som sammen kalles megaloblastiske anemier.
Artikkelindeks
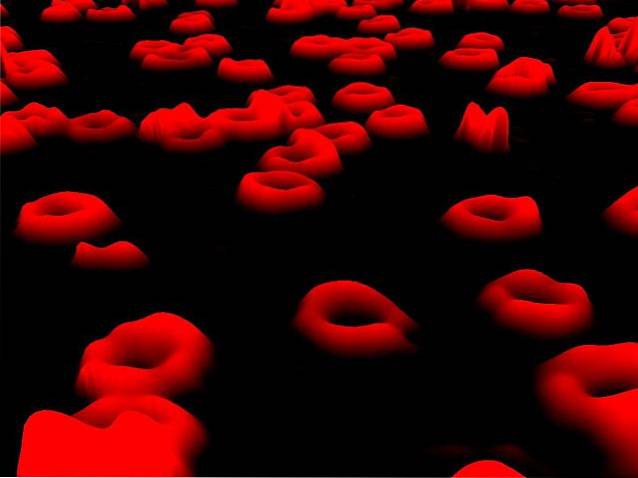
De har en karakteristisk morfologi som ligner på bikonkave plater, og deres hovedfunksjon er å utføre oksygentransport (O2) til kroppens forskjellige vev, samtidig som det avgifter dem fra karbondioksid (CO2) som produseres under cellulær åndedrett..
Denne utvekslingen av CO2 mot O2 er mulig fordi disse cellene huser store mengder av et karakteristisk rødt protein kalt hemoglobin, som er i stand til å samhandle med begge kjemiske arter gjennom en hemgruppe som er tilstede i strukturen..
En egenart hos disse cellene hos pattedyr med hensyn til resten av virveldyr er mangelen på kjerne og cytoplasmatiske organeller. Imidlertid har det i løpet av de innledende produksjonsfasene i de tidlige stadiene av embryonal utvikling blitt observert at de cellulære forløperne som de stammer fra, utgjør en overgående kjerne.
Sistnevnte er ikke overraskende gitt at de tidlige stadiene av embryoutviklingen vanligvis er like i alle virveldyr, og divergerer bare trinnene som kompromitterer større differensiering..
Erytroblaster er celler som vil gi opphav til modne erytrocytter etter å ha gjennomgått påfølgende hendelser med celledifferensiering..
Disse forløpercellene stammer fra en vanlig myeloid stamfar i benmargen til virveldyr som kjernefysiske celler, forsynt med kjerner og cellulære organeller..
Endringer i innholdet av cytoplasmaet og i omorganiseringen av cytoskelettet vil kulminere i generasjonen av erytrocytter som er klare til sirkulasjon. Disse endringene reagerer på miljøstimuli som indikerer reduksjon av oksygen i vevet og derfor et behov i produksjonen av erytrocytter..
Erytropoiesis er betegnelsen som brukes til å definere prosessen der produksjon og utvikling av røde blodlegemer foregår, nødvendig for å opprettholde oksygentilførselen til de forskjellige organer og vev..
Denne prosessen er finregulert av virkningen av erytropoietin (EPO), et nyresyntesehormon som igjen moduleres av oksygenkonsentrasjonene som er tilgjengelige i vevet..
Oksygenkonsentrasjoner med lavt vev induserer EPO-syntese ved hypoksiinducerbar transkripsjonsfaktor (HIF-1), som stimulerer erytrocyttproliferasjon ved binding til EpoR-reseptorer, tilstede på erytrocyttforløperceller.
Hos pattedyr utføres erytropoiesis i to trinn som kalles primitiv erytropoiesis og definitiv erytropoiesis..
Den første forekommer i plommesekken under embryonal utvikling, noe som gir opphav til store kjernefysiske erytroblaster, mens den andre forekommer i fosterleveren og fortsetter i beinmargen etter den andre svangerskapsmåneden, og genererer mindre enukleare erytrocytter..
Andre proteiner som det antipoptotiske cytokinet Bcl-X, hvis transkripsjon er regulert av transkripsjonsfaktoren GATA-1, påvirker også erytropoieseprosessen positivt. I tillegg er tilførsel av jern, vitamin B12 og folsyre også nødvendig.
I prosessen med endelig erytropoies dannes erytrocytter i benmargen fra en udifferensiert stamcelle eller vanlig myeloid stamfar som er i stand til å gi opphav til andre celler som granulocytter, monocytter og blodplater..
Denne cellen må motta passende ekstracellulære signaler for å kompromittere dens differensiering mot erytroidavstamningen..
Når denne forpliktelsen er oppnådd, begynner en sekvens av differensieringshendelser som begynner med dannelsen av pronormoblasten, også kjent som proerythroblast. En stor erytroblast forløpercelle med en kjerne.
Deretter vil proerythroblast oppleve en progressiv reduksjon i kjernevolumet ledsaget av en økning i hemoglobinsyntese. Alle disse endringene skjer sakte når denne cellen passerer gjennom forskjellige celletrinn: basofil erytroblast eller normoblast, polykromatisk erytroblast og ortokromatisk erytroblast.
Prosessen avsluttes med det totale tapet av kjernen, så vel som organellene som er tilstede i den ortokromatiske erytroblasten, med opprinnelse til en moden erytrocyt..
For å endelig nå dette, må sistnevnte passere gjennom retikulocyttstadiet, en enukleert celle som fremdeles inneholder organeller og ribosomer i cytoplasmaet. Fullstendig fjerning av kjernen og organellene utføres av eksocytose.
Modne erytrocytter forlater benmargen i blodstrømmen der de forblir i sirkulasjon i omtrent 120 dager, før de blir oppslukt av makrofager. Derfor er erytropoiesis en prosess som skjer kontinuerlig gjennom en organismes liv..
Når erytoblaster utvikler seg mot fullstendig differensiering til en moden erytrocytt, gjennomgår de flere endringer i cytoskelettet, så vel som i uttrykket av celleadhesjonsproteiner..
Actin-mikrofilamenter depolymeriserer og et nytt spektrinbasert cytoskjelett monteres. Spectrin er et perifert membranprotein som ligger på det cytoplasmatiske ansiktet som interagerer med ankyrin, et protein som formidler bindingen av cytoskjelettet med Band 3 transmembranproteinet.
Disse endringene i cytoskelettet og i uttrykket av Epo-reseptorer, så vel som mekanismene som modulerer dem, er kritiske for erytroid modning..
Dette skyldes det faktum at de formidler etableringen av interaksjoner mellom erytroblaster og celler som er tilstede i benmargsmikromiljøet, og letter overføring av de nødvendige signalene for å begynne og avslutte differensiering..
Når differensieringen er fullført, oppstår nye endringer som favoriserer tapet av vedheft av cellene til marg og frigjøres i blodet der de vil utføre sin funksjon..
Feil under differensiering av erytroblaster i benmargen gir opphav til utseende av blodpatologier, for eksempel megaloblastiske anemier. Disse stammer fra mangler i tilførselen av vitamin B12 og folat som er nødvendig for å fremme erytroblastdifferensiering..
Begrepet megaloblastisk refererer til den store størrelsen som erytroblaster og til og med erytrocytter når som et produkt av ineffektiv erytropoies, preget av mangelfull DNA-syntese..



Ingen har kommentert denne artikkelen ennå.