De sphingomyelin det er den mest utbredte sfingolipiden i dyrevev: dens tilstedeværelse er funnet i alle cellemembraner som hittil er undersøkt. Den har strukturelle likheter med fosfatidylkolin når det gjelder polarhodegruppen, og det er derfor det også er klassifisert som et fosfolipid (fosfosfingolipid).
På 1880-tallet isolerte forsker Johann Thudichum en eterløselig lipidkomponent fra hjernevev og kalte den sfingomyelin. Senere, i 1927, ble strukturen til denne sfingolipiden rapportert som N-acyl-sfingosin-1-fosfokolin.
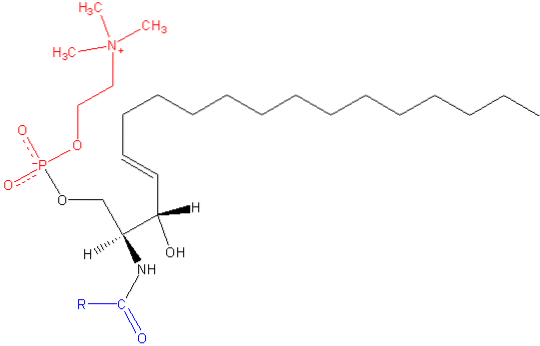
Som de andre sfingolipidene, har sfingomyelin både strukturelle og cellesignalerende funksjoner, og er spesielt rikelig i nervesvev, spesielt i myelin, en kappe som dekker og isolerer aksonene til visse nevroner.
Distribusjonen av den er studert gjennom subcellulær fraksjonering og enzymatisk nedbrytningseksperimenter med sfingomyelinaser, og resultatene indikerer at mer enn halvparten av sfingomyelin i eukaryote celler finnes i plasmamembranen. Dette avhenger imidlertid av celletypen. I fibroblaster utgjør det for eksempel nesten 90% av totale lipider.
Dysregulering av syntesen og metabolismen av dette lipidet fører til utvikling av komplekse patologier eller lipidose. Et eksempel på disse er arvelig Niemann-Pick sykdom, preget av hepatosplenomegali og progressiv nevrologisk dysfunksjon.
Artikkelindeks
Sphingomyelin er et amfipatisk molekyl som består av et polarhode og to apolare haler. Polarhodegruppen er et fosfokolinmolekyl, så det kan se ut som glyserofosfolipidfosfatidylkolin (PC). Imidlertid er det vesentlige forskjeller med hensyn til grensesnittet og den hydrofobe regionen mellom disse to molekylene..
Den vanligste basen i et sphingomyelinmolekyl fra pattedyr er ceramid, sammensatt av sfingosin (1,3-dihydroksy-2-amino-4-oktadecen), som har en dobbeltbinding i trans mellom karbonene i posisjon 4 og 5 i hydrokarbonkjeden. Det mettede derivatet, sfinganin, er også vanlig, men finnes i mindre grad.
Lengden på de hydrofobe halene til sfingomyelin varierer fra 16 til 24 karbonatomer, og fettsyresammensetningen varierer avhengig av vevet..
Sfingomyeliner av den hvite substansen i den menneskelige hjerne har for eksempel nervonsyre, de av grå materie inneholder hovedsakelig stearinsyre, og den utbredte formen i blodplater er arakidonat..
Det er generelt en forskjell i lengde mellom de to fettsyrekjedene til sfingomyelin, som synes å favorisere "interdigitering" fenomener mellom hydrokarboner i motsatte monolag. Dette gir membranen spesiell stabilitet og spesielle egenskaper sammenlignet med andre membraner som er dårligere i denne sfingolipiden..
I grensesnittområdet av molekylet har sfingomyelin en amidgruppe og et fritt hydroksyl ved karbon 3, som kan tjene som givere og akseptorer av hydrogenbindinger for intra- og intermolekylære bindinger, viktig i definisjonen av sidedomener og interaksjon. Med forskjellige typer molekyler.
Produktene av sfingosinmetabolisme -ceramid, sfingosin, sfingosin 1-fosfat og diacylglyserol- er viktige cellulære effektorer og gir det en rolle i flere cellulære funksjoner, som apoptose, utvikling og aldring, cellesignalering, blant andre..
Takket være den tredimensjonale "sylindriske" strukturen til sfingomyelin, kan dette lipidet danne mer kompakte og ordnede membrandomener, noe som har viktige funksjonelle implikasjoner fra proteinets synspunkt, siden det kan etablere spesifikke domener for noen integrerte membranproteiner..
Lipidflåter, membranfaser eller ordnede mikrodomener av sfingolipider som sfingomyelin, noen glyserofosfolipider og kolesterol, representerer stabile plattformer for assosiasjon av membranproteiner med forskjellige funksjoner (reseptorer, transportører, etc.).
Caveolae er invaginasjoner av plasmamembranen som rekrutterer proteiner med GPI-ankre og er også rike på sfingomyelin.
Kolesterol, på grunn av sin strukturelle stivhet, påvirker strukturen til cellemembraner betydelig, spesielt i forhold relatert til fluiditet, og det er derfor det regnes som et essensielt element.
Fordi sfingomyeliner har både hydrogenbindingsdonorer og akseptorer, antas det at de er i stand til å danne mer “stabile” interaksjoner med kolesterolmolekyler. Det er derfor det sies at det er en positiv sammenheng mellom nivåene av kolesterol og sfingomyelin i membranene..
Syntesen av sfingomyelin forekommer i Golgi-komplekset, der ceramidet som transporteres fra endoplasmatisk retikulum (ER), modifiseres ved overføring av et fosfokolinmolekyl fra fosfatidylkolin, med samtidig frigjøring av et diacylglyserolmolekyl. Reaksjonen katalyseres av SM-syntase (ceramid: fosfatidylkolin fosfokolin transferase).
Det er også en annen vei for sfingomyelinproduksjon som kan oppstå ved å overføre en fosfoetanolamin fra fosfatidyletanolamin (PE) til ceramid, med påfølgende fosfoetanolaminmetylering. Dette antas å være spesielt viktig i noen PE-rike nervevev..
Sfingomyelinsyntase finnes på luminal side av Golgi-kompleksmembranen, noe som er i samsvar med den ekstra cytoplasmiske plasseringen av sfingomyelin i de fleste celler.
På grunn av egenskapene til den polare gruppen av sfingomyelin og det tilsynelatende fraværet av spesifikke translokaser, avhenger den topologiske orienteringen av dette lipidet av enzymsyntasen..
Sfingomyelin-nedbrytning kan forekomme i både plasmamembranen og lysosomene. Lysosomal hydrolyse til ceramid og fosfokolin avhenger av sur sfingomyelinase, et løselig lysosomalt glykoprotein hvis aktivitet har en optimal pH på rundt 4,5.
Hydrolyse i plasmamembranen katalyseres av en sfingomyelinase som fungerer ved pH 7,4 og som krever toverdige magnesium- eller manganioner for å fungere. Andre enzymer som er involvert i metabolismen og resirkuleringen av sfingomyelin, finnes i forskjellige organeller som kobles til hverandre gjennom vesikulære transportveier..



Ingen har kommentert denne artikkelen ennå.