De peripatric spesiering, i evolusjonær biologi refererer det til dannelsen av nye arter fra et lite antall individer som ble isolert i periferien til denne opprinnelige populasjonen.
Det ble foreslått av Ernst Mayr og er en av hans mest kontroversielle teorier i evolusjon. Opprinnelig ble det kalt spesiering etter grunnleggereffekt, for senere å bli kalt parapatrisk spesiering.
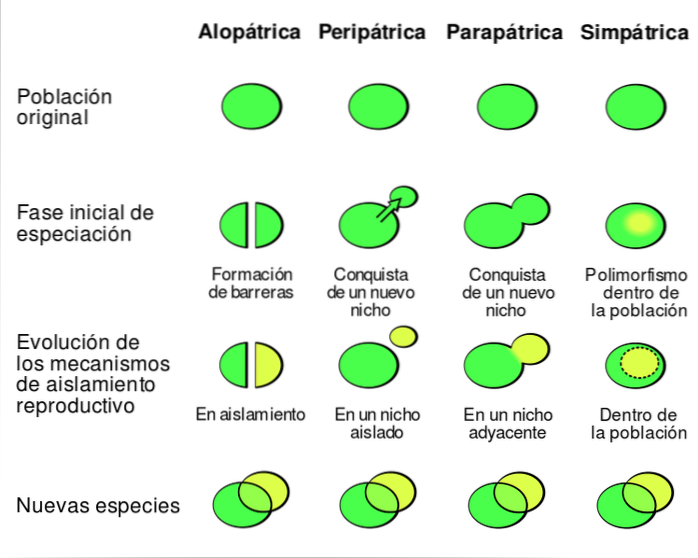
Den nye arten oppstår ved grensene for den sentrale befolkningen, som har et større antall individer. Under spesifikasjonsprosessen kan flyten mellom populasjoner reduseres til det maksimale til den slutter å eksistere. Dermed utgjør perifer befolkning med tiden en ny art.
I denne spesieringsmodellen skiller dispersjonen og koloniseringsfenomenene seg ut. Når individer sprer seg, utsettes de for selektive trykk (for eksempel miljøforhold) som er forskjellige fra den opprinnelige populasjonen som til slutt fører til divergens..
Genetisk drift ser ut til å ha en spesiell rolle i den parapatriske spesieringsmodellen, siden den isolerte populasjonen generelt er liten og stokastiske faktorer har større effekt i populasjoner med reduserte størrelser..
Artikkelindeks
I følge Curtis & Schnek (2006) er peripatrisk spesiering definert som “en gruppe individer grunnlegger en ny befolkning. Hvis den grunnleggende gruppen er liten, kan den ha en spesiell genetisk konfigurasjon, ikke representativ for den opprinnelige befolkningen. ".
Dette kan skje hvis befolkningen opplever en flaskehals (en betydelig reduksjon i antall individer) eller hvis et lite antall individer migrerer til periferien. Disse migrantene kan bestå av et enkelt par eller en enkelt inseminert kvinne.
Det samme kan skje når en befolkning opplever en nedgang i størrelse. Når denne reduksjonen oppstår, reduseres fordelingsområdet etter tur, og små isolerte populasjoner forblir i periferien til den opprinnelige befolkningen. Genstrømning mellom disse gruppene er ekstremt lav eller null.
Denne mekanismen ble foreslått av evolusjonsbiologen og ornitologen Ernst Mayr, på midten av 1950-tallet.
Ifølge Mayr begynner prosessen med spredning av en liten gruppe. På et tidspunkt (Mayr forklarer ikke klart hvordan det skjer, men tilfeldighet spiller en avgjørende rolle) stopper migrasjonen mellom den opprinnelige befolkningen og den lille isolerte befolkningen..
Mayr beskrev denne modellen i en artikkel som fokuserte på studien av fugler fra New Guinea. Teorien var basert på perifere fuglepopulasjoner som skiller seg sterkt fra tilstøtende populasjoner. Mayr aksepterer at hans forslag stort sett er spekulativt.
En annen innflytelsesrik biolog i evolusjonsteorier, Hennig, aksepterte denne mekanismen og kalte den kolonisering..
Etter klassifiseringen av Curtis & Schnek (2006) av spesifikasjonsmekanismene som er foreslått av disse forfatterne, er det tre hovedmodeller for spesiering ved divergens: allopatrisk, parapatrisk og sympatrisk. Mens de øyeblikkelige spesieringsmodellene er peripatric og polyploidy speciering.
Futuyma (2005) plasserer derimot parapatrisk spesiering som en type allopatrisk spesiering - sammen med vikarians. Derfor klassifiseres peripatrisk spesiering i henhold til opprinnelsen til den reproduktive barrieren..
Mayr foreslår at den genetiske endringen av den isolerte populasjonen skjer raskt, og genstrømmen med den opprinnelige populasjonen blir avskåret. I følge denne forskerens begrunnelse ville allelfrekvensene på noen steder være forskjellige fra de i den opprinnelige populasjonen, bare på grunn av prøvetakingsfeil - med andre ord genetisk drift..
Samplingsfeilen er definert som tilfeldige avvik mellom det som teoretisk er forventet og de oppnådde resultatene. Anta for eksempel at vi har en pose med røde og svarte bønner i forholdet 50:50. Ved en ren tilfeldighet, når jeg velger 10 bønner fra posen, kan jeg få 4 røde og 6 svarte.
Ekstrapolerer dette didaktiske eksemplet til populasjoner, kan det være at "grunnlegger" -gruppen som vil bli etablert i periferien ikke har de samme allelfrekvensene som den opprinnelige populasjonen..
Mayrs hypotese innebærer en betydelig evolusjonær endring som skjer raskt. Videre, ettersom den geografiske plasseringen er ganske spesifikk og begrenset, kombinert med tidsfaktoren, ville den ikke bli dokumentert i fossilregisteret..
Denne påstanden prøver å forklare det plutselige utseendet til arten i fossilregisteret, uten de forventede mellomstadiene. Derfor forutsa Mayrs ideer teorien om punktert likevekt, foreslått av Gould og Eldredge i 1972..
Ikke alle levende organismer virker potensielle kandidater for peripatrisk spesiering for å produsere en forandring i deres populasjoner.
Enkelte egenskaper, som liten spredningskapasitet og et mer eller mindre stillesittende liv, gjør at noen linjer er utsatt for grupper slik at denne spesieringsmodellen kan handle på dem. I tillegg må organismer ha en tendens til å strukturere seg i små populasjoner..
I den hawaiiske øygruppen er den dannet av en rekke øyer og atoller bebodd av et stort antall endemiske arter.
Skjærgården har trukket evolusjonsbiologers oppmerksomhet for de nesten 500 artene (noen endemiske) av slekten. Drosophila som bor på øyene. Det foreslås at den enorme diversifiseringen av gruppen skjedde takket være koloniseringen av noen få individer på nærliggende øyer.
Denne hypotesen er bekreftet ved å anvende molekylære teknikker på disse hawaiiske befolkningene..
Undersøkelser har avdekket at de nært beslektede artene finnes på nærliggende øyer, og at arter som nylig har gått fra hverandre, bor på de nye øyene. Disse fakta støtter ideen om en peripatrisk spesiering.
Lizard av arten Uta stansburiana Den tilhører familien Phrynosomatidae og er innfødt i USA og Nord-Mexico. Blant de mest fremragende egenskapene er eksistensen av polymorfier i befolkningen.
Disse populasjonene representerer et godt eksempel på peripatrisk spesiering. Det er en befolkning som bor på øyene i Gulf of California og varierer mye sammenlignet med sine kolleger i USA..
Islandindivider skiller seg vidt i forskjellige egenskaper som størrelse, fargelegging og økologiske vaner..



Ingen har kommentert denne artikkelen ennå.