
De postmendeliansk arv Den inkluderer alle ideene og / eller verkene som oppstod etter studiene publisert av den religiøse naturforskeren Gregorio Mendel i 1865, og som forklarer genetiske fenomener eller atferd som er forskjellig fra de som er beskrevet av dette.
Gregorio Mendel regnes som faren til genetikk, siden han ved å studere kryss mellom erteplanter etablerte de første arvelovene for mer enn 100 år siden. Den første loven, kalt "lov om segregering", foreslår at gener kan eksistere i forskjellige eller alternative former.

Disse alternative formene kalles "alleler" og de påvirker direkte de fenotypiske egenskapene til en art. Alleler kan bestemme hårfarge, øyenfarge, hårtype osv..
I sin første lov foreslo Mendel at hvert fenotypisk trekk som overføres fra foreldre til barna deres kontrolleres av to alleler, som hver kommer fra en forelder (en fra "moren" og en fra "faren").
I følge dette, når foreldrene bidrar med forskjellige alleler, resulterer avkommet i en blanding, der en av allelene er dominerende (den som uttrykkes i fenotypen) og den andre er recessiv (den blir ikke sett, eller er "latent" ")..
Mendels andre lov er kjent som "Loven om uavhengig arv", siden Mendel i sine kors observerte at arvemønsteret som karakteriserte allelene som definerte et trekk, ikke påvirket arvemønsteret til allelene som definerte til et annet trekk.
Artikkelindeks
Siden Gregor Mendel foreslo sine berømte lover for mer enn 100 år siden, har det blitt belyst nye mekanismer for å forklare arven til tegn, hvorav mange kan sies å være i strid med eller skille seg betydelig fra de som er beskrevet av Mendel..
Et eksempel på fenomenene som er beskrevet av "moderne" vitenskap, hvis fremskritt bare har vært mulig takket være utviklingen av eksperimentelle strategier og forskningsmetoder, er arv knyttet til sex, ufullstendig dominans og kodominans, blant andre..
Kjønnskoblet arv har å gjøre med gener som bare finnes på kjønnskromosomene. Hos mennesker bestemmes sex av X- og Y-kromosomene. Kvinner har to eksemplarer av X (XX) og menn har en av X og en av Y (XY)..
Genene som finnes på hver av disse to kromosomene er de som bidrar til de egenskapene som er knyttet til sex. Gener på X-kromosomet er tilstede hos menn og kvinner, men Y-gener er bare hos menn.
Kjønnsbundet arv ble observert og beskrevet for første gang av William Bateson, en av de første forskerne som “gjenoppdaget” Mendels arbeid og utnyttet dem til å utføre nye eksperimenter basert på hans konklusjoner..
Bateson laget eksperimentelle kors av høner med fjærdrakt i forskjellige farger, noen med mørke og glatte fjær og andre med glatte fjær og med stripete mønster..
Gjennom disse korsene observerte han at hos avkomene hadde de resulterende hannene alltid fjærdrakt med stripete mønstre.
Først i tredje generasjon, da en stripet hann ble krysset med en høne med mørke og glatte fjær, ble det oppnådd noen haner med svart fjærdrakt. Denne resulterende andelen av korsene trosset proporsjonene foreslått av Mendel.
Etter Batesons arbeid fant Thomas Henry Morgan en logisk forklaring på de rare resultatene Bateson oppnådde, og tilskrev forskjellen i fargelegging hos kyllinger til det faktum at det stripete mønsteret ble kodet av en dominerende allel som ble funnet på X-kromosomet..
Hos høner, i motsetning til hos mennesker, tilsvarer det heterogametiske kjønnet (XY) kvinnene, mens det homogametiske kjønnet tilsvarer hannene (XX).
Allelen studert av Bateson og Morgan ble da uttrykt av alle hannene med stripete fjærdrakt, men når en hann arvet to X-kromosomer med allelen som tilsvarte den svarte fargen, så dukket hannene med mørk fjærdrakt og ikke noe stripet mønster opp..
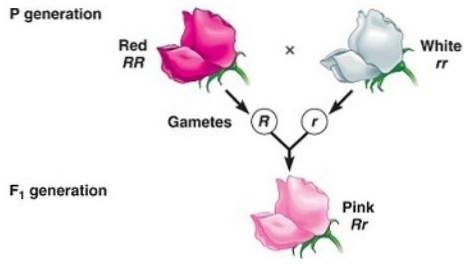
Ufullstendig dominans er når fenotypen observert hos avkommet som skyldes kryssing av to homozygote individer (den ene dominerende og den andre recessive) er en "mellomliggende" fenotype mellom den dominerende og den recessive..
Det har å gjøre med den "dominerende" allelens "manglende evne" til å fullstendig maskere uttrykket til den recessive allelen, og det er derfor den resulterende fenotypen tilsvarer en blanding av egenskapene kodet av begge alleler, mange ganger beskrevet som en "fortynnet fenotype ”Av den dominerende.
For eksempel i Dondiego-plantene om natten (Mirabilis jalapa) den typiske fargen på villblomsterblader er rød. Når en plante med røde kronblader krysses med en ren linje plante med hvite kronblader, oppnås det at alle avkom er rosa (rosa) kronblader.
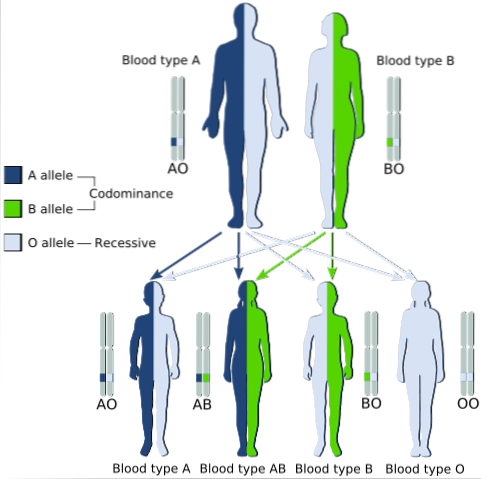
Når to dominerende homozygote individer krysses og en tredje fenotype observeres i avkommet som tilsvarer de to egenskapene som er bestemt av de to foreldrenes alleler, snakker vi om kodominans.
I dette tilfellet, fortsetter med et eksempel som ligner på blomstene i forrige eksempel, vil den "tredje fenotypen" ikke svare til rosa blomster, men til en plante med hvite blomster og røde blomster på samme stamme, siden de to allelene "codominate", og ingen av dem maskerer den andre.
Mange ganger er kodominans og ufullstendig dominans imidlertid vanskelig å skille fenotypisk, siden begge produserer en fenotype av et individ som ser ut til å uttrykke to forskjellige alleler..
Imidlertid er individene som presenterer kodominante alleler genotypisk forskjellige fra de som presenterer ufullstendig dominans, siden i det ene tilfellet uttrykkes de to dominerende allelene i samme mengde, og i det andre er proporsjonene ulik..
Et klassisk eksempel på kodominans er typen blodgruppe hos mennesker, siden alleler som koder for antigener A og B i menneskelige populasjoner er kodominante.
Det vil si at når alleler A og B blir funnet sammen i cellene til et individ, har han blodtype AB.
De er recessive alleler for noe tegn som, når de finnes i homozygot form, er dødelige for individet. Individer som presenterer disse allelene, blir ikke født, siden de forhindrer normal funksjon av molekylmaskineriet og forhindrer oversettelse av gener.
Mange steder i genomet til et individ samhandler med andre deler av det samme genomet, det vil si at den spesifikke modifiseringen av en allel i genomet kan direkte betinget ekspresjon av andre alleler i cellene til et individ..
Disse interaksjonene er vanskelige å bestemme og klassifiseres i to forskjellige typer: gener som samhandler i samme genetiske vei og de som samhandler på forskjellige genetiske veier..
Et klassisk og velkjent eksempel på gener som samhandler i forskjellige genetiske veier, har å gjøre med genene som bestemmer hudfargen til maisslanger (Pantherophis guttatus), hvis vanlige farge er et gjentatt mønster av svarte og oransje flekker.

Denne fenotypen er produsert av to forskjellige pigmenter som er under streng genetisk kontroll. Et gen bestemmer den oransje fargen og den har to alleler: en O +, som koder for pigmentet og en annen O-, som bestemmer det totale fraværet av det oransje pigmentet..
Et annet forskjellig gen bestemmer tilstedeværelsen eller fraværet av det svarte pigmentet, allelene til dette genet er kjent som B +, den som bestemmer tilstedeværelsen av det svarte pigmentet og B-, som bestemmer fraværet av pigmentet.
Begge gener er ikke koblet og bestemmer uavhengig fargen på slanger, det vil si at en slange med O + / B + genotypen er en normal slange med en oransje og svart fenotype, men enhver genotypisk variasjon vil resultere i en annen fenotype..
Det klassiske eksemplet på disse genene er det som er beskrevet for "bellflower" -blomstene (av Campanulaceae-familien), hvis "ville" fenotype tilsvarer planter med blå blomster..
Den blå fargen på blåklokkeblomster bestemmes ikke av et gen, men av en metabolsk vei der flere enzymer, hver kodet av et annet gen, deltar i syntesen av et pigment..
Dermed er det fenotyper preget av å være et resultat av samspillet mellom flere gener, hvis produkter deltar i den samme metabolske veien, og hvor mutasjonen eller feilfunksjonen til noen av disse gir en fenotype, i dette tilfellet, av hvite, depigmenterte blomster..

Ingen har kommentert denne artikkelen ennå.