De histoner er grunnleggende proteiner som samhandler med DNA for dannelse av nukleosomer, som utgjør kromatintrådene som utgjør kromosomer i eukaryote organismer.
Nukleosomer, komplekser som består av DNA og proteiner, ble oppdaget i 1974, og det er histoner som samler dette basale nivået av kromatinorganisasjon. Imidlertid har eksistensen av histonproteiner vært kjent siden før 1960-tallet..
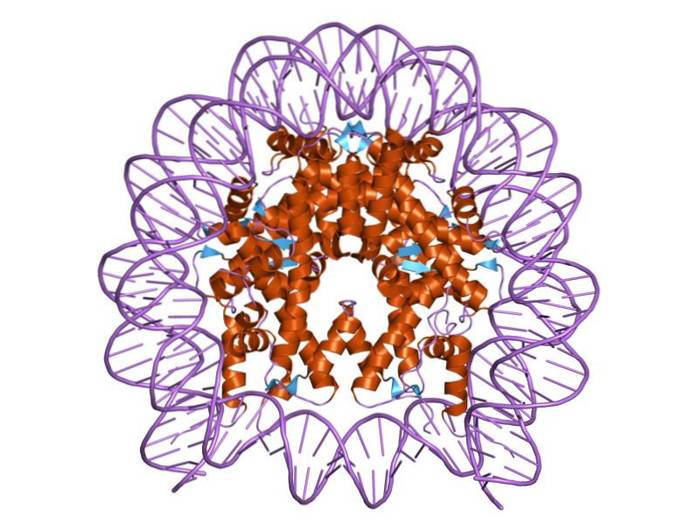
Histoner er organisert på en slik måte at dobbeltbånd-DNAet brytes rundt et proteinsenter som består av disse proteinene som samhandler tett med hverandre. Histonsenteret er formet som en plate, og DNA går rundt 1,7 ganger rundt det..
Flere hydrogenbindinger tillater DNA å binde seg til proteinsenteret dannet av histonene i hvert nukleosom. Disse bindingene er for det meste dannet mellom aminosyre-ryggraden i histoner og sukker-fosfat-ryggraden i DNA. Noen hydrofobe interaksjoner og ioniske bindinger deltar også.
Proteiner kjent som "kromatin remodeling complexes" er ansvarlige for nedbryting og dannelse av kryssbindingene mellom DNA og histoner, slik at transkripsjonsmaskineriet kan komme inn i DNA som finnes i nukleosomene..
Til tross for nærheten til nukleinsyrer til proteinsenteret som dannes av histoner, er disse ordnet på en slik måte at de, om nødvendig, tillater innføring av transkripsjonsfaktorer og andre proteiner relatert til genuttrykk eller gendemping..
Histoner kan gjennomgå forskjellige modifikasjoner som genererer flere varianter, noe som muliggjør eksistensen av mange forskjellige former for kromatin som har egenskapen til å modulere genetisk uttrykk på forskjellige måter..
Artikkelindeks
De er et av de mest konserverte eukaryote proteinene i naturen. For eksempel har ertehiston H4 vist seg å variere i bare to av de 102 aminosyreposisjonene til ku H4-proteinet..
Histoner er relativt små proteiner, med ikke mer enn 140 aminosyrer. De er rike på basiske aminosyrerester, så de har en netto positiv ladning, noe som bidrar til deres interaksjon med den negativt ladede nukleinsyren for å danne nukleosomer..
Nukleosomale og brodannende histoner er kjent. De nukleosomale histonene er H3, H4, H2A og H2B, mens bindingshistonene tilhører familien til histoner H1.
Under nukleosommontering dannes opprinnelig spesifikke dimerer H3-H4 og H2A-H2B. To H3-H4-dimerer går deretter sammen for å danne tetramerer som senere kombineres med H2A-H2B-dimerer og danner det oktameriske sentrum..
Alle histoner syntetiseres hovedsakelig i løpet av S-fasen i cellesyklusen, og nukleosomene samles i de spirende DNA-spiralene, like etter replikasjonsgaffelen..
Den generelle strukturen til histoner inkluderer en basisk aminosyreregion og en globulær karboksylregion som er sterkt konservert blant eukaryote organismer..
Et strukturelt motiv kjent som "histonfold", sammensatt av tre alfa-helixer forbundet med to hårnål og danner et lite hydrofobt senter, er ansvarlig for protein-protein-interaksjonen mellom histonene som utgjør nukleosomet..
Det er denne folden av histoner som utgjør det kuleformede karboksyldomenet til disse nukleosomale proteiner i alle eukaryoter..
Histoner har også små "haler" eller aminoterminale og andre karboksylterminale regioner (tilgjengelige for proteaser), ikke mer enn 40 aminosyrer lange. Begge regionene er rike på basiske aminosyrer som kan gjennomgå flere post-translationelle kovalente modifikasjoner..
I eukaryoter er det to familier av bindende histoner, differensiert fra hverandre etter deres struktur. Noen har en trepartsstruktur, med det globulære domenet beskrevet ovenfor flankert av "ustrukturerte" N- og C-terminale domener; mens andre bare har et C-terminal domene.
Selv om de fleste histoner er konservert, kan spesifikke varianter oppstå under embryogenese eller modning av spesialiserte celler i noen organismer. Noen strukturelle variasjoner har å gjøre med endringer etter translasjon, for eksempel følgende:
-Fosforylering: det antas å være relatert til modifisering av graden av kondensering av kromatin, og det forekommer ofte i serinrester.
-Acetylering: assosiert med kromosomale regioner som er transkripsjonelt aktive. Det forekommer normalt på sidekjedene til lysinrester. Når de forekommer på disse restene, reduseres deres positive ladning og reduserer dermed affiniteten til proteiner for DNA..
-Metylering: kan forekomme som mono-, di- eller trimetylering av lysinrester som stikker ut fra proteinkjernen.
Spesifikke enzymer er ansvarlige for disse kovalente modifikasjonene i histoner. Disse enzymene inkluderer histon-acetyltransferaser (HAT), histondeacetylasekomplekser (HDAC), og histon-metyltransferaser og demetylaser..
Karakteriseringen av histonene har blitt utført ved forskjellige biokjemiske teknikker, blant hvilke kromatografiene som er basert på svake kationbytterharpikser skiller seg ut..
Enkelte forfattere etablerer en form for klassifisering der fem hovedtyper histoner skilles ut i eukaryoter: FI, med 21 kDa proteiner; F2A1 eller FIV, på pluss eller minus 11,3 kDa; F2A2 eller FIIbI, 14,5 kDa; F2B eller FIIb2, med en molekylvekt på 13,7 kDa, og F3 eller FIII, på 15,3 kDa.
Alle disse typer histoner, med unntak av IF-gruppen, finnes i ekvimolære mengder i celler..
En annen klassifisering, med samme gyldighet og kanskje den mest brukte i dag, foreslår at det eksisterer to forskjellige typer histoner, nemlig: de som er en del av nukleosomets oktamer og bindings- eller brohistonene, som forbinder nukleosomene mellom Ja.
Noen varianter kan også forekomme mellom arter, og i motsetning til kjernehistoner syntetiseres varianter under mellomfase og settes inn i det forhåndsdannede kromatinet gjennom en prosess avhengig av energien som frigjøres fra ATP-hydrolyse..
Sentrum av et nukleosom består av et par av hver av de fire bestanddelene histoner: H2a, H2b, H3 og H4; på hvilke DNA-segmenter på ca. 145 basepar er viklet.
Histoner H4 og H2B er i prinsippet uforanderlige. Noen variasjoner er imidlertid tydelige i histonene H3 og H2A, hvis biofysiske og biokjemiske egenskaper endrer nukleosomets normale natur..
En variant av histon H2A hos mennesker, H2A.Z-proteinet har en stor sur region og kan fremme nukleosomstabilitet avhengig av histon H3-variantene som det er assosiert med..
Disse histonene viser en viss variasjon mellom arter, et spesielt tilfelle er det for histon H2B, der den første tredjedelen av molekylet er svært variabel..
Bindings- eller brodannende histoner er klasse H1-histoner. Disse er ansvarlige for foreningen mellom nukleosomer og beskyttelsen av DNA som stikker ut i begynnelsen og på slutten av hver partikkel..
I motsetning til nukleosomale histoner, har ikke alle histoner av H1-typen den kuleformede regionen av "folden" av histon. Disse proteinene binder seg til DNA mellom nukleosomene, og letter transkripsjonsmessig et skifte i kromatinbalanse mot en mer kondensert og mindre aktiv tilstand..
Studier har knyttet disse histonene til aldring, DNA-reparasjon og apoptotiske prosesser, og det er derfor de antas å spille en avgjørende rolle for å opprettholde genomisk integritet.
Alle aminosyrerester av histoner deltar på en eller annen måte i deres interaksjon med DNA, noe som forklarer det faktum at de er så konserverte blant kongedømmene til eukaryote organismer.
Deltakelse av histoner i emballasjen av DNA i form av kromatin er av stor relevans for komplekse flercellede organismer der de forskjellige cellelinjene bare kan spesialisere seg ved å endre tilgjengeligheten av deres gener til transkripsjonsmaskineriet..
Transkripsjonelt aktive genomregioner er tette i nukleosomer, noe som tyder på at assosiasjonen av DNA med histonproteiner er avgjørende for den negative eller positive reguleringen av transkripsjonen..
Gjennom en celles levetid avhenger responsen på et stort antall stimuli, både interne og eksterne, av små endringer i kromatin, som normalt har å gjøre med ombygging og post-translationell modifisering av histonene som finnes i nært forhold til DNA.
Flere histonvariabler tjener forskjellige funksjoner i eukaryoter. En av dem har å gjøre med deltakelse av en variant av histon H3 i dannelsen av de sentromere strukturer som er ansvarlige for segregering av kromosomer under mitose..
Motstykket til dette proteinet i andre eukaryoter har vist seg å være viktig for montering av proteinkinetokoren som spindelmikrotubuli binder seg under mitose og meiose..



Ingen har kommentert denne artikkelen ennå.