Mitokondrier er karakteristiske intracellulære organeller av alle eukaryote celler. De har ansvaret for en viktig del av cellulær energimetabolisme og er hovedstedet for ATP-produksjon i celler med aerob metabolisme..
Sett under mikroskopet, er disse organellene like store som en bakterie og deler mange av deres genetiske egenskaper med prokaryoter, for eksempel tilstedeværelsen av et sirkulært genom, bakterieribosomer og overførings-RNA som ligner på de andre prokaryoter..

Endosymbiotisk teori foreslår at disse organellene oppsto i eukaryote stamfedre for millioner av år siden fra prokaryote celler som "parasitiserte" primitive eukaryoter, og ga dem muligheten til å leve i aerobiose og bruke oksygen til energi, motta ly i retur. Og næringsstoffer..
Siden genomet deres må ha blitt redusert, ble dannelsen av disse organellene i stor grad avhengig av import av proteiner som syntetiseres i cytosolen fra gener kodet i kjernen, også fra fosfolipider og andre metabolitter, som ble tilpasset komplekse transportmaskiner.
I dag er det kjent at mitokondriene fungerer som "kraftkildene" til alle aerobe eukaryote celler, og at Krebs-syklusen forekommer i dem, syntesen av pyrimidiner, aminosyrer og noen fosfolipider. I sin indre forekommer oksidasjon av fettsyrer også, hvorfra store mengder ATP oppnås..
Som i alle cellulære organismer, er mitokondrie-DNA utsatt for mutasjoner, noe som resulterer i mitokondriale dysfunksjoner som fører til nevrodegenerative lidelser, kardiomyopatier, metabolske syndromer, kreft, døvhet, blindhet og andre patologier..
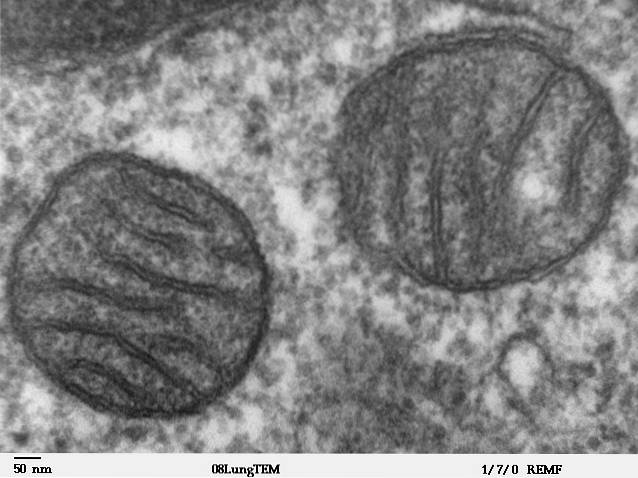
Mitokondrier er ganske store cytosoliske organeller, deres størrelse overstiger størrelsen på kjernen, vakuolene og kloroplaster i mange celler; volumet kan representere opptil 25% av det totale volumet av cellen. De har en karakteristisk orm- eller pølselignende form og kan være flere mikrometer lange..
De er organeller omgitt av en dobbel membran som har sitt eget genom, det vil si at det er et DNA-molekyl fremmed (annerledes) enn DNA som finnes i cellekjernen. De har også ribosomalt RNA og overfører RNA av seg selv..
Til tross for det ovennevnte, er de avhengige av nukleære gener for produksjon av de fleste av deres proteiner, som er spesielt merket under deres oversettelse i cytosolen som skal transporteres til mitokondriene..
Mitokondriene deler seg og formerer seg uavhengig av celler; deres oppdeling skjer ved mitose, noe som resulterer i dannelsen av en mer eller mindre nøyaktig kopi av hver enkelt. Med andre ord, når disse organellene deler seg, gjør de det ved å "dele seg i to".
Antall mitokondrier i eukaryote celler er sterkt avhengig av celletypen og dens funksjon; det vil si i samme vev av en flercellet organisme, noen celler kan ha et større antall mitokondrier enn andre. Et eksempel på dette er hjertemuskelcellene, som har et stort antall mitokondrier.
Mitokondrier er essensielle organeller for aerobe celler. Disse fungerer i integrasjonen av mellommetabolisme i flere metabolske veier, blant hvilke oksidativ fosforylering for produksjon av ATP i celler skiller seg ut..
Innvendig forekommer oksidasjon av fettsyrer, Krebs-syklusen eller trikarboksylsyrene, urea-syklusen, ketogenesen og glukoneogenesen. Mitokondrier spiller også en rolle i syntesen av pyrimidiner og noen fosfolipider.
De er også involvert, delvis, i metabolismen av aminosyrer og lipider, i syntesen av hemgruppen, i kalsiumhomeostase og i prosessene med programmert celledød eller apoptose.
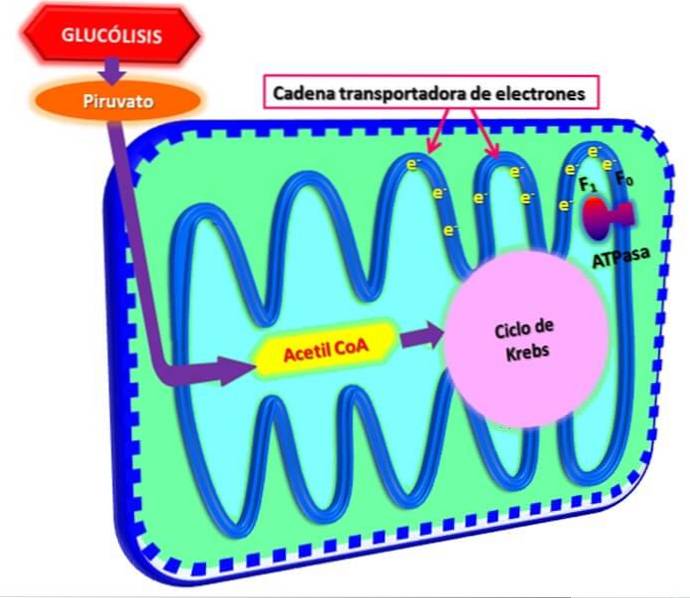
Glykolyse, prosessen med oksiderende glukose for å utvinne energi fra den i form av ATP, forekommer i det cytosoliske rommet. I celler med aerob metabolisme, pyruvat (sluttproduktet av glykolytisk vei per se) transporteres til mitokondriene, der den tjener som et substrat for pyruvatdehydrogenase-enzymkomplekset.
Dette komplekset er ansvarlig for dekarboksylering av pyruvat til C02, NADH og acetyl-CoA. Det sies at energien i denne prosessen er "lagret" i form av acetyl-CoA-molekyler, siden disse er de som "kommer inn" i Krebs-syklusen, hvor deres acetyldel er fullstendig oksidert til CO2 og vann..
På samme måte oksyderes lipidene som sirkulerer gjennom blodstrømmen og kommer inn i cellene direkte i mitokondriene gjennom en prosess som begynner ved karbonylenden av dem, og hvorved to karbonatomer elimineres samtidig i hver "retur", danner ett acetyl-CoA-molekyl av gangen.
Nedbrytningen av fettsyrer ender med produksjonen av NADH og FADH2, som er molekyler med høyenergielektroner som deltar i oksidasjonsreduksjonsreaksjoner..
I løpet av Krebs-syklusen blir CO elimin eliminert som avfallsprodukt, i mellomtiden transporteres NADH- og FADH2-molekylene til elektrontransportkjeden i den indre membranen i mitokondriene, hvor de brukes i den oksidative fosforyleringsprosessen..
Enzymer som deltar i elektrontransportkjeden og oksidativ fosforylering finnes i mitokondriens indre membran. I denne prosessen tjener NADH- og FADH2-molekylene som "transportører" av elektroner når de overfører dem fra de oksiderende molekylene til transportkjeden..
Disse elektronene frigjør energi når de passerer gjennom transportkjeden, og denne energien brukes til å kaste ut protoner (H +) fra matrisen inn i mellomrommet gjennom den indre membranen, og generere en protongradient..
Denne gradienten fungerer som en energikilde som er koblet til andre reaksjoner som krever energi, for eksempel generering av ATP ved fosforylering av ADP..
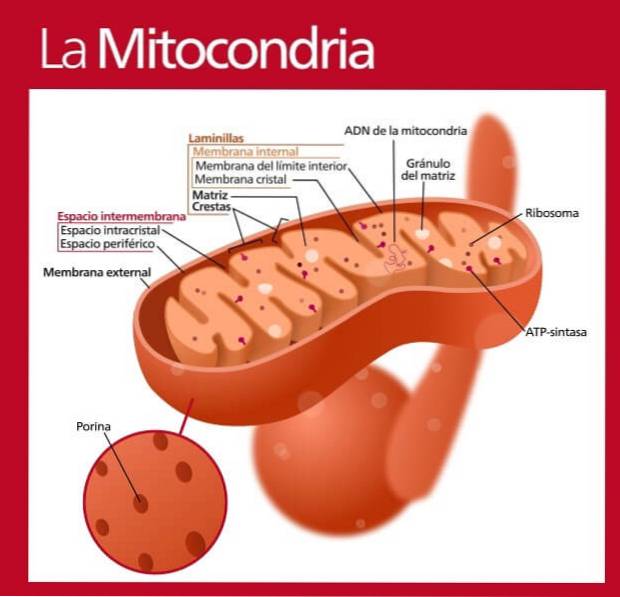
Disse organellene er unike blant andre cytosoliske organeller av flere grunner, noe som kan forstås ut fra kunnskap om deres deler..
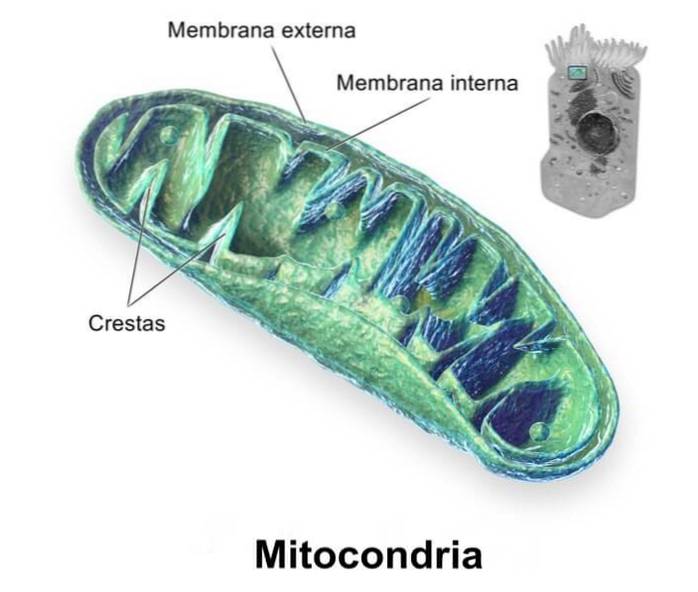
Mitokondrier er, som allerede nevnt, cytosoliske organeller omgitt av en dobbel membran. Denne membranen er delt inn i den ytre mitokondrie-membranen og den indre mitokondrie-membranen, veldig forskjellig fra hverandre og atskilt fra hverandre av intermembranrommet..
Denne membranen er den som fungerer som grensesnittet mellom cytosol og mitokondrie lumen. Som alle biologiske membraner, er den ytre mitokondrie membranen et lipid dobbeltlag som perifere og integrerte proteiner er knyttet til.
Mange forfattere er enige om at forholdet mellom proteiner og lipider i denne membranen er nær 50:50, og at denne membranen er veldig lik den for gramnegative bakterier..
Proteinene til den ytre membranen fungerer i transport av forskjellige typer molekyler mot intermembranrommet, mange av disse proteinene er kjent som "poriner", siden de danner kanaler eller porer som tillater fri passering av små molekyler fra den ene siden til annet. annet.
Denne membranen inneholder et veldig stort antall proteiner (nesten 80%), mye større enn det for den ytre membranen og en av de høyeste prosentene i hele cellen (det høyeste protein: lipidforholdet).
Det er en membran som er mindre gjennomtrengelig for passering av molekyler og danner flere bretter eller rygger som rager ut mot lumen eller mitokondriell matrise, selv om antallet og arrangementet av disse foldene varierer betydelig fra en type celle til en annen, selv i samme organisme..
Den indre mitokondrielle membranen er det viktigste funksjonelle rommet til disse organellene, og dette skyldes i hovedsak deres tilknyttede proteiner.
Dens bretter eller rygger spiller en spesiell rolle i å øke membranoverflaten, noe som gir et rimelig bidrag til å øke antall proteiner og enzymer som deltar i mitokondriefunksjoner, det vil si i oksidativ fosforylering, hovedsakelig (elektrontransportkjede)..
Som det kan utledes av navnet, er intermembranrommet det som skiller de ytre og indre mitokondrielle membranene..
Siden den ytre mitokondrielle membranen har mange porer og kanaler som letter fri diffusjon av molekyler fra den ene siden til den andre, har intermembranrommet en sammensetning som ligner den på cytosolen, i det minste med hensyn til ioner og visse molekyler. liten i størrelse.
Den mitokondrielle matrisen er mitokondriens indre rom og er stedet der mitokondrie genomisk DNA blir funnet. I tillegg er det i denne "væsken" også noen av de viktige enzymene som deltar i cellulær energimetabolisme (mengden proteiner er større enn 50%).
I den mitokondrielle matrisen er det for eksempel enzymer som tilhører Krebs-syklusen eller trikarboksylsyresyklusen, som er en av hovedveiene for oksidativ metabolisme i aerobe organismer eller celler..
Mitokondrier er unike cytosoliske organeller i celler, siden de har sitt eget genom, det vil si at de har sitt eget genetiske system, som er forskjellig fra cellenes (innesluttet i kjernen).
Genomet til mitokondrier består av sirkulære DNA-molekyler (slik som prokaryoter), hvorav det kan være flere kopier per mitokondrion. Størrelsen på hvert genom avhenger mye av arten som vurderes, men hos mennesker er dette for eksempel omtrent 16 kb.
Genene som koder for noen mitokondrieproteiner, finnes i disse DNA-molekylene. Det er også gener som koder for ribosomale RNAer og overfører RNAer som er nødvendige for oversettelse av proteinene kodet av mitokondriegenomet inne i disse organellene..
Den genetiske koden som brukes av mitokondrier for å "lese" og "oversette" proteiner som er kodet i genomet, er noe forskjellig fra den universelle genetiske koden.
Menneskelige mitokondrielle sykdommer er en ganske heterogen gruppe sykdommer, siden de har å gjøre med mutasjoner i både mitokondrie og atom DNA..
Avhengig av typen mutasjon eller genetisk defekt, er det forskjellige patologiske manifestasjoner relatert til mitokondriene, som kan påvirke ethvert organsystem i kroppen og mennesker i alle aldre.
Disse mitokondriale feilene kan overføres fra en generasjon til en annen gjennom moderens rute, gjennom X-kromosomet eller gjennom den autosomale ruten. Av denne grunn er mitokondrieforstyrrelser virkelig heterogene både i det kliniske aspektet og i de vevsspesifikke manifestasjonene..
Blant noen av de kliniske manifestasjonene relatert til mitokondriefeil er:
Dyreceller og planteceller inneholder mitokondrier. I begge celletyper utfører disse organellene likeverdige funksjoner, og selv om de ikke er veldig viktige, er det noen små forskjeller mellom disse organellene..
De viktigste forskjellene mellom dyre- og plantemitokondrier har å gjøre med morfologi, størrelse og noen genomiske egenskaper. Dermed kan mitokondrier variere i størrelse, antall, form og organisering av de indre ryggene; selv om dette også gjelder for de forskjellige celletyper i samme organisme.
Størrelsen på mitokondriegenomet hos dyr er litt mindre enn plantene (henholdsvis k 20kb vs 200kb). I motsetning til dyremitokondrier koder de i planteceller for tre typer ribosomalt RNA (dyr koder bare for to).
Imidlertid er plantemitokondrier avhengig av noe kjernefysisk overførings-RNA for syntesen av proteinene deres..
I tillegg til de som allerede er nevnt, er det ikke mange andre forskjeller mellom mitokondriene i dyreceller og planteceller, som rapportert av Cowdry i 1917..


Ingen har kommentert denne artikkelen ennå.