
De handlingspotensial Det er et kortvarig elektrisk eller kjemisk fenomen som oppstår i hjernens nerveceller. Det kan sies at det er meldingen som et nevron overfører til andre nevroner.
Handlingspotensialet produseres i cellekroppen (kjernen), også kalt soma. Den beveger seg gjennom hele aksonet (en forlengelse av nevronet, som en ledning) til den når slutten, kalt terminalknappen.
Handlingspotensialer på et gitt axon har alltid samme varighet og intensitet. Hvis aksonen forgrener seg til andre prosesser, deles handlingspotensialet, men intensiteten reduseres ikke.
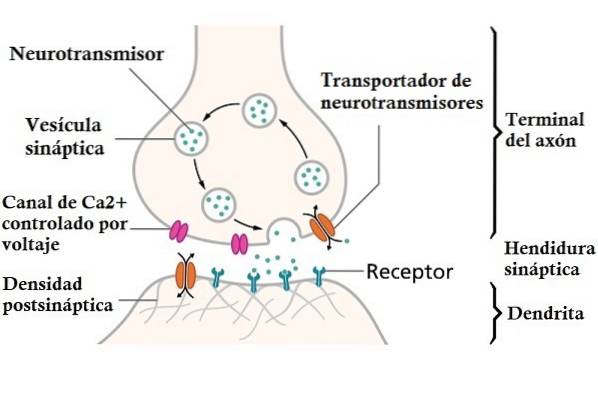
Når handlingspotensialet når terminalens knapper i nevronet, skiller de ut kjemikalier som kalles nevrotransmittere. Disse stoffene begeistrer eller hemmer nevronet som mottar dem, og er i stand til å generere et handlingspotensial i nevronet.
Mye av det som er kjent om neurons handlingspotensialer kommer fra eksperimenter med gigantiske blekksprutaksoner. Det er lett å studere på grunn av størrelsen, siden den strekker seg fra hode til hale. De tjener slik at dyret kan bevege seg.
Artikkelindeks
Nevroner har en annen elektrisk ladning innvendig enn utenfor. Denne forskjellen kalles Membranpotensial.
Når et nevron er i hvilepotensial, betyr at den elektriske ladningen ikke endres av eksiterende eller hemmende synaptiske potensialer.
På den annen side, når andre potensialer påvirker det, kan membranpotensialet reduseres. Dette er kjent som depolarisering.
Tvert imot, når membranpotensialet øker med hensyn til sitt normale potensial, kalles et fenomen hyperpolarisering.
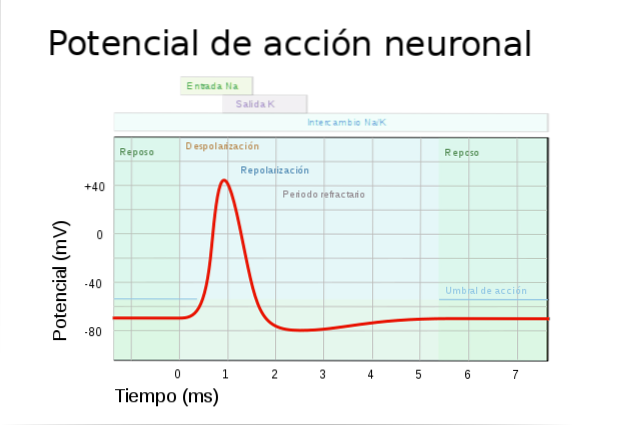
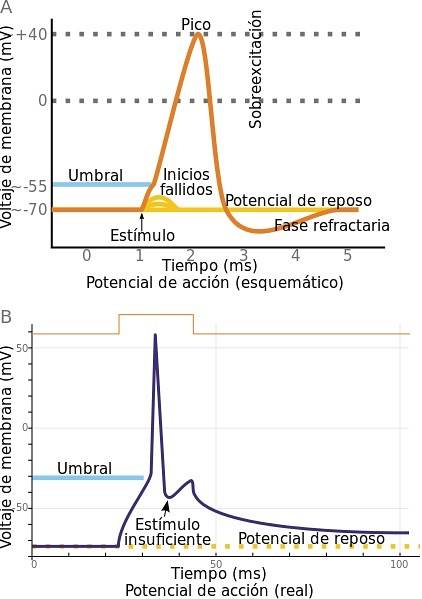
Når en veldig rask reversering av membranpotensialet oppstår plutselig, er det en handlingspotensial. Dette består av en kort elektrisk impuls, som oversettes til meldingen som beveger seg gjennom neuronets akson. Det begynner i cellehuset og når terminalknappene.
Det er viktig at for at et handlingspotensial skal skje, må elektriske endringer nå en terskel, kalt eksitasjonsterskel. Det er verdien av membranpotensialet som nødvendigvis må nås for at handlingspotensialet skal oppstå.
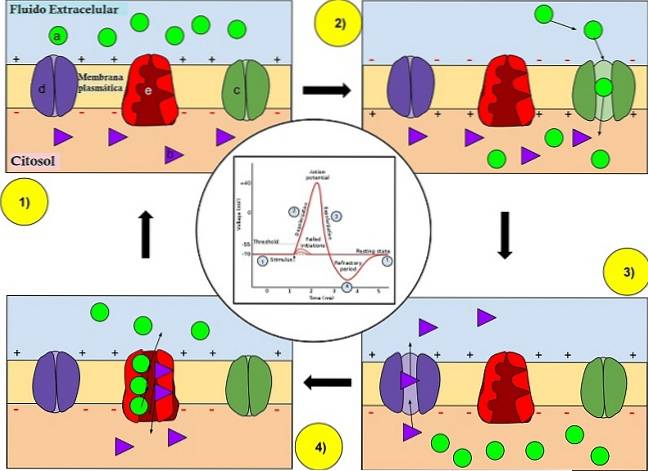
Under normale forhold er nevronen klar til å motta natrium (Na +) inni. Imidlertid er membranen ikke veldig permeabel for dette ionet..
I tillegg har de velkjente "natrium-kalium-transportørene" et protein som finnes i cellemembranen som er ansvarlig for å fjerne natriumioner fra den og innføre kaliumioner i den. Nærmere bestemt introduserer den for hver 3 natriumioner den tre kaliumioner.
Disse transportørene holder natriumnivået lavt i cellen. Hvis permeabiliteten til cellen skulle øke og mer natrium kom inn i den plutselig, ville membranpotensialet endre seg radikalt. Tilsynelatende er det dette som utløser et handlingspotensial.
Spesielt vil permeabiliteten til membranen for natrium øke, når disse kommer inn i nevronet. Samtidig som kaliumioner kan komme ut av cellen.
Celler har innebygd mange proteiner som kalles i membranen ionekanaler. Disse har åpninger der ioner kan komme inn i eller forlate celler, selv om de ikke alltid er åpne. Kanaler stenges eller åpnes basert på visse hendelser.
Det finnes flere typer ionekanaler, og hver og en er vanligvis spesialisert for å utføre bestemte typer ioner utelukkende.
For eksempel kan en åpen natriumkanal passere mer enn 100 millioner ioner per sekund..
Nevroner overfører informasjon elektrokjemisk. Dette betyr at kjemikalier produserer elektriske signaler.
Disse kjemikaliene har en elektrisk ladning, og det er derfor de kalles ioner. Det viktigste i nervesystemet er natrium og kalium, som har en positiv ladning. I tillegg til kalsium (2 positive ladninger) og klor (en negativ ladning).
Det første trinnet for at et handlingspotensial skal skje er en endring i cellens membranpotensial. Denne endringen må overskride eksitasjonsterskelen.
Spesielt er det en reduksjon i membranpotensialet, som kalles depolarisering..
Som en konsekvens åpnes natriumkanaler innebygd i membranen, slik at natrium kan trenge inn i nevronen. Disse drives av diffusjonskrefter og elektrostatisk trykk.
Siden natriumioner er positivt ladede, gir de en rask endring i membranpotensialet.
Axonmembranen har både natrium- og kaliumkanaler. Sistnevnte åpnes imidlertid senere, fordi de er mindre følsomme. Det vil si at de trenger et høyere nivå av depolarisering for å åpne, og det er derfor de åpner senere.
Det kommer en tid da handlingspotensialet når sin maksimale verdi. Fra denne perioden blokkeres og lukkes natriumkanalene.
De vil ikke lenger kunne åpne igjen før membranen når sitt hvilepotensial igjen. Som et resultat vil ikke mer natrium kunne komme inn i nevronet..
Kaliumkanalene forblir imidlertid åpne. Dette gjør at kaliumioner kan strømme gjennom cellen..
På grunn av diffusjon og elektrostatisk trykk, da det indre av aksonet er positivt ladet, skyves kaliumionene ut av cellen. Dermed gjenvinner membranpotensialet sin vanlige verdi. Litt etter litt stenger kaliumkanalene.
Denne utgangen av kationer fører til at membranpotensialet gjenoppretter sin normale verdi. Når dette skjer, begynner kaliumkanalene å lukkes igjen..
Så snart membranpotensialet når sin normale verdi, lukkes kaliumkanalene helt. Noe senere reaktiveres natriumkanalene som forberedelse for en annen depolarisering for å åpne dem.
Til slutt, skiller natrium-kaliumtransportørene ut natriumet som hadde kommet inn og gjenvunnet kaliumet som tidligere hadde gått.
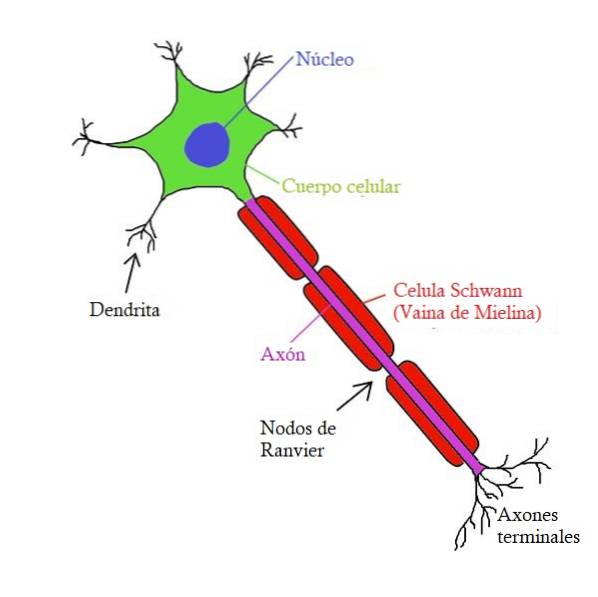
Aksonen består av en del av nevronet, en kabellignende forlengelse av nevronet. De kan være for lange til at nevroner som er fysisk langt fra hverandre, kan koble til og sende informasjon til hverandre..
Handlingspotensialet forplanter seg langs aksonen og når terminalknappene for å sende meldinger til neste celle. Hvis vi målte intensiteten til handlingspotensialet fra forskjellige områder av aksonen, ville vi finne at dens intensitet forblir den samme i alle områder.
Dette skjer fordi aksonal ledning følger en grunnleggende lov: loven om alt eller ingenting. Det vil si at et handlingspotensial er gitt eller ikke. Når den begynner, beveger den seg gjennom aksonen til slutten, og holder alltid samme størrelse, den øker eller reduseres ikke. Hva mer, hvis et axon forgrener seg, splittes handlingspotensialet, men opprettholder størrelsen..
Handlingspotensialer starter på slutten av aksonen som er festet til nevronens soma. De reiser vanligvis bare i én retning.
Du lurer kanskje på dette punktet: Hvis handlingspotensialet er en alt eller ingenting prosess, hvordan oppstår visse atferd som muskelsammentrekning som kan variere mellom forskjellige intensitetsnivåer? Dette skjer etter loven om frekvens.
Det som skjer er at et enkelt handlingspotensial ikke gir informasjon direkte. I stedet bestemmes informasjonen av utslippsfrekvensen eller skytefrekvensen til et akson. Det vil si frekvensen der handlingspotensial oppstår. Dette er kjent som "frekvensloven"..
Dermed vil en høy frekvens av handlingspotensialer gi en veldig intens muskelsammentrekning..
Det samme er tilfelle med persepsjon. For eksempel må en veldig lys visuell stimulus, som skal fanges, gi en høy "avfyringshastighet" i aksonene festet til øynene. På denne måten gjenspeiler frekvensen av handlingspotensialene intensiteten til en fysisk stimulans..
Derfor kompletteres loven om alt eller ingenting med frekvensloven.
Handlingspotensialer er ikke de eneste klassene av elektriske signaler som forekommer i nevroner. For eksempel, når du sender informasjon gjennom en synaps, blir det gitt en liten elektrisk impuls i nevronens membran som mottar dataene..
Noen ganger kan en svak depolarisering som er for svak til å produsere et handlingspotensial, endre membranpotensialet..
Imidlertid reduseres denne endringen gradvis når den beveger seg gjennom aksonen. I denne typen informasjonsoverføring åpner eller lukkes verken natrium- eller kaliumkanalene..
Dermed fungerer aksonen som en sjøkabel. Når signalet overføres gjennom det, reduseres amplituden. Dette er kjent som nedadgående ledning, og det oppstår på grunn av aksonens egenskaper..
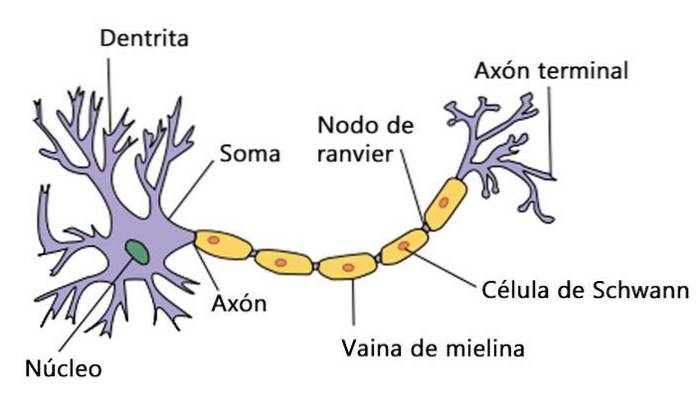
Aksonene til nesten alle pattedyr er dekket av myelin. Det vil si at de har segmenter omgitt av et stoff som tillater nerveledning, noe som gjør det raskere. Myelin brytes rundt aksonen uten at ekstracellulær væske når den.
Myelin produseres i sentralnervesystemet av celler kalt oligodendrocytter. Mens det i det perifere nervesystemet produseres av Schwann-celler.
Myelin-segmentene, kjent som myelin-kapper, er delt fra hverandre ved bare områder av aksonen. Disse områdene kalles Ranviers knuter og de er i kontakt med ekstracellulær væske..
Handlingspotensialet overføres annerledes i et umyelinisert akson (som ikke er dekket av myelin) enn i et myelinisert.
Handlingspotensialet kan vandre gjennom den myelinbelagte aksonale membranen på grunn av ledningens egenskaper. Aksonen leder på denne måten den elektriske endringen fra stedet der handlingspotensialet oppstår til neste node på Ranvier..
Denne endringen avtar litt, men er sterk nok til å forårsake et handlingspotensial i neste knute. Dette potensialet blir deretter utløst eller gjentatt i hver knute av Ranvier, og transporterer seg gjennom det myeliniserte området til neste knute..
Denne typen ledning av handlingspotensialer kalles saltdannelse. Navnet kommer fra det latinske "saltare", som betyr "å danse". Konseptet er fordi impulsen ser ut til å hoppe fra node til node.
Denne typen kjøring har sine fordeler. Først og fremst for å spare energi. Natrium-kaliumtransportører bruker mye energi på å trekke overflødig natrium fra innsiden av aksonen under handlingspotensialer.
Disse natrium-kaliumtransportørene er lokalisert i områdene av axonet som ikke er dekket av myelin. Imidlertid kan natrium i et myelinisert akson bare komme inn i nodene til Ranvier. Derfor kommer mye mindre natrium inn, og på grunn av dette må mindre natrium pumpes ut, så natrium-kaliumtransportørene må jobbe mindre.
En annen fordel med myelin er hastighet. Et handlingspotensial blir ført raskere i et myelinisert akson, siden impulsen "hopper" fra en node til en annen, uten å måtte gå gjennom hele axonen.
Denne økningen i hastighet får dyr til å tenke og reagere raskere. Andre levende vesener, som blekksprut, har axoner uten myelin som får fart på grunn av en økning i størrelsen. Blekksprutaksoner har en stor diameter (ca. 500 µm), som gjør at de kan bevege seg raskere (ca. 35 meter per sekund).
Imidlertid beveger aksjonspotensialene seg i samme hastighet i aksonene til katter, selv om disse har en diameter på bare 6 µm. Det som skjer er at disse aksonene inneholder myelin.
Et myelinisert akson kan lede aksjonspotensialer med en hastighet på ca. 432 kilometer i timen, med en diameter på 20 µm..

Ingen har kommentert denne artikkelen ennå.