De prometafase det er et stadium i celledelingsprosessen, mellom mellom profase og metafase. Det er preget av samspillet mellom delende kromosomer og mikrotubuli som er ansvarlige for å skille dem. Prometaphase forekommer i både mitose og meiose, men med forskjellige egenskaper.
Det klare målet med all celledeling er å produsere flere celler. For å oppnå dette må cellen opprinnelig duplisere DNA-innholdet; repliker det. I tillegg til dette må cellen skille disse kromosomene på en slik måte at det spesielle formålet med hver deling av cytoplasma blir oppfylt..

I mitose er det samme antall kromosomer fra morscellen i dattercellene. I meiose I, skillet mellom homologe kromosomer. I meiose II, skillet mellom søsterkromatider. Det vil si at du ved slutten av prosessen får de fire forventede meiotiske produktene.
Cellen håndterer denne kompliserte mekanismen gjennom bruk av spesialiserte komponenter som mikrotubuli. Disse er organisert av sentrosomet i de fleste eukaryoter. I andre, tvert imot, for eksempel høyere planter, fungerer en annen type organisasjonssenter for mikrotubuli.
Artikkelindeks

Mikrotubuli er lineære polymerer av proteinet tubulin. De er involvert i nesten alle cellulære prosesser som involverer forskyvning av noen indre struktur. De er en integrert del av cytoskelettet, cilia og flagella.
Når det gjelder planteceller, spiller de også en rolle i den interne strukturelle organisasjonen. I disse cellene danner mikrotubuli en slags veggteppe festet til plasmamembranens indre overflate..
Denne strukturen, som kontrollerer inndeling av planter, er kjent som den kortikale organisasjonen av mikrotubuli. På tidspunktet for mitotisk deling kollapser de for eksempel til en sentral ring som vil være det fremtidige stedet for sentralplaten, i planet der cellen vil dele seg..
Mikrotubuli består av alfa-tubulin og beta-tubulin. Disse to underenhetene danner en heterodimer, som er den grunnleggende strukturelle enheten av tubulinfilamenter. Polymeriseringen av dimerer fører til dannelse av 13 protofilamenter i en lateral organisasjon som gir opphav til en hul sylinder.

De hule sylindrene med denne strukturen er mikrotubuli, som ved sin egen sammensetning viser polaritet. Det vil si at den ene enden kan vokse ved tilsetning av heterodimerer, mens den andre kan gjennomgå subtraksjoner. I sistnevnte tilfelle krymper mikrotubuli, i stedet for å forlenge seg i den retningen.
Mikrotubuli kjerner (det vil si begynner å polymerisere) og organiseres i mikrotubuli-organiseringssentre (COM). COM er assosiert med sentrosomer under inndeling i dyreceller.
I høyere planter, som ikke har sentrosomer, er COM til stede på analoge steder, men består av andre komponenter. I cilia og flagella er COM plassert i basal retning mot motorstrukturen.
Forskyvningen av kromosomer under celledeling oppnås gjennom mikrorør. Disse formidler den fysiske interaksjonen mellom kromosomens sentromer og COM.
Ved målrettede depolymeriseringsreaksjoner vil metafasekromosomer til slutt bevege seg mot polene i delende celler..
Korrekt mitotisk kromosomadskillelse er en som garanterer at hver dattercelle mottar et komplement av kromosomer som er identiske med modercellens..
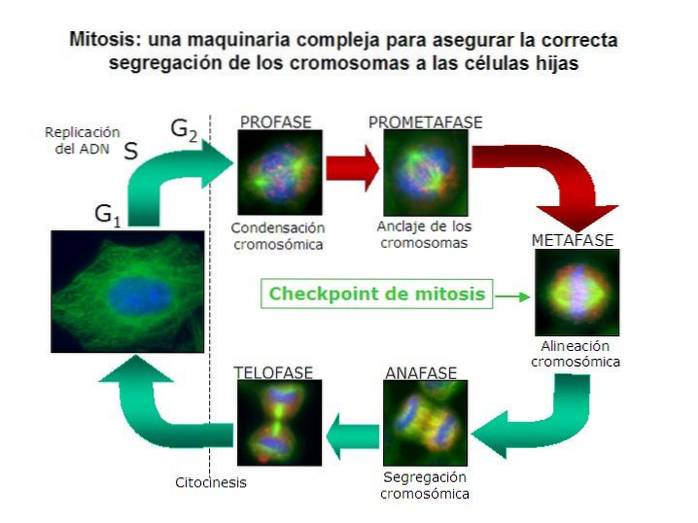
Dette betyr at cellen må skille hvert par dupliserte kromosomer i to separate, individuelle kromosomer. Det vil si at den må adskille søsterkromatidene til hvert homologe par fra hele komplementet av stamcellens kromosomer..
I åpne mitoser er prosessen med forsvinden av kjernekapslingen kjennetegnet på prometafase. Dette gjør at den eneste hindringen mellom MOC og kromosomens sentromer forsvinner..
Fra MOC-er polymeriseres lange mikrotubuli-filamenter og forlenges mot kromosomene. Ved å finne et sentromer opphører polymerisasjonen og et kromosom festet til en COM oppnås..
I mitose er kromosomene doble. Derfor er det også to sentromerer, men fortsatt forenet i samme struktur. Dette betyr at på slutten av mikrotubuli-polymeriseringsprosessen vil vi ha to av dem per duplisert kromosom..
Én glødetråd vil feste en sentromer til en COM, og en annen til søsterkromatiden festet til COM motsatt den første..
I lukkede mitoser er prosessen nesten identisk med den forrige, men med stor forskjell; kjernekonvolutten forsvinner ikke. Derfor er COM internt og er assosiert med den interne kjernekonvolutten gjennom den kjernefysiske lamellen..
I semi-lukkede (eller semi-åpne) mitoser forsvinner kjernekonvolutten bare på de to motsatte punktene der en mitotisk COM eksisterer utenfor kjernen..
Dette betyr at mikrotubuli i disse mitosene trenger inn i kjernen for å kunne mobilisere kromosomene i trinn etter prometafasen..
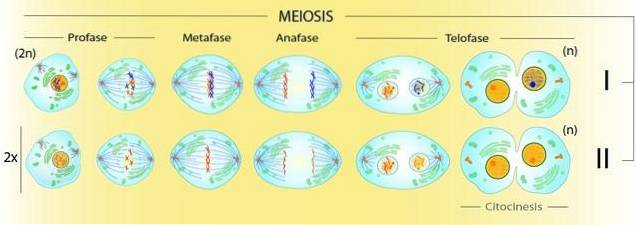
Ettersom meiose involverer produksjon av fire 'n' celler fra en '2n' celle, må det være to divisjoner av cytoplasma. La oss se på det slik: på slutten av metafase I vil det være fire ganger flere kromatider enn sentromerer synlige under mikroskopet.
Etter første deling vil det være to celler med dobbelt så mange kromatider som sentromerer. Først på slutten av den andre cytoplasmatiske inndelingen vil alle sentromerer og kromatider bli individualisert. Det vil være like mange sentromerer som det er kromosomer.
Nøkkelproteinet for at disse komplekse interkromatininteraksjonene skal finne sted i mitose og meiose er cohesin. Men det er flere komplikasjoner i meiose enn i mitose. Det er derfor ikke overraskende at meiotisk kohesin er forskjellig fra mitotisk.
Kohesiner tillater kohesjon av kromosomer under deres mitotiske og meiotiske kondensasjonsprosess. I tillegg tillater og regulerer de samspillet mellom søsterkromatider i begge prosesser..
Men i meiose fremmer de også noe som ikke forekommer i mitose: homolog parring, og de påfølgende synapsene. Disse proteinene er forskjellige i hvert tilfelle. Vi kan si at meiose uten en kohesin som skiller den, ikke ville være mulig.
Mekanisk sett er sentromere / COM-interaksjonen den samme i all celledeling. Imidlertid vil cellen ikke skille søsterkromatider i prometafase I av meiose I, slik den gjør i mitose..
I kontrast har den meiotiske tetrad fire kromatider i et tilsynelatende dobbelt sett med sentromerer. I denne strukturen er det noe annet som ikke er tilstede i mitose: chiasmata.
Kiasmer, som er fysiske knutepunkter mellom homologe kromosomer, er det som skiller hvilke sentromerer som må skilles: de av homologe kromosomer.
I prometafase I blir det således dannet forbindelser mellom sentromerene i homologene og COM på motsatte poler av cellen..
Denne prometaphase II er mer lik mitotisk prometaphase enn meiotisk prometaphase I. I dette tilfellet vil COM-ene "lansere" mikrotubuli på dupliserte sentromerer av søsterkromatider..
Dermed vil to celler bli produsert med individuelle kromosomer, produktet av en kromatid fra hvert par. Derfor vil celler med det haploide kromosomale komplementet arten bli gitt.


Ingen har kommentert denne artikkelen ennå.