De DNA-emballasje er et begrep som definerer den kontrollerte komprimeringen av DNA i cellen. I ingen celle (og faktisk ikke engang i virus) er DNA-fri, løs og i sann løsning.
DNA er et ekstremt langt molekyl som også alltid samhandler med et stort utvalg av forskjellige proteiner. For prosessering, nedarving og kontroll av uttrykket av genene det bærer, vedtar DNA en bestemt romlig organisasjon. Dette oppnås av cellen ved å strengt kontrollere hvert trinn av DNA-pakking på forskjellige komprimeringsnivåer..
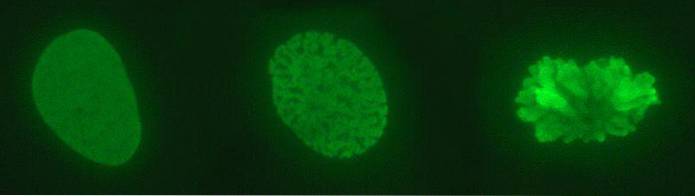
Virus har forskjellige pakningsstrategier for nukleinsyrene. En av favorittene er dannelsen av kompakte spiraler. Det kan sies at virus er nukleinsyrer pakket i sine egne proteiner som dekker, beskytter og mobiliserer dem.
I prokaryoter er DNA assosiert med proteiner som bestemmer dannelsen av komplekse sløyfer i en struktur som kalles nukleoid. Det maksimale nivået av DNA-komprimering i en eukaryot celle er derimot mitotisk eller meiotisk kromosom..
Det eneste tilfellet der et B-DNA ikke er pakket er et forskningslaboratorium som forfølger dette formålet..
Artikkelindeks
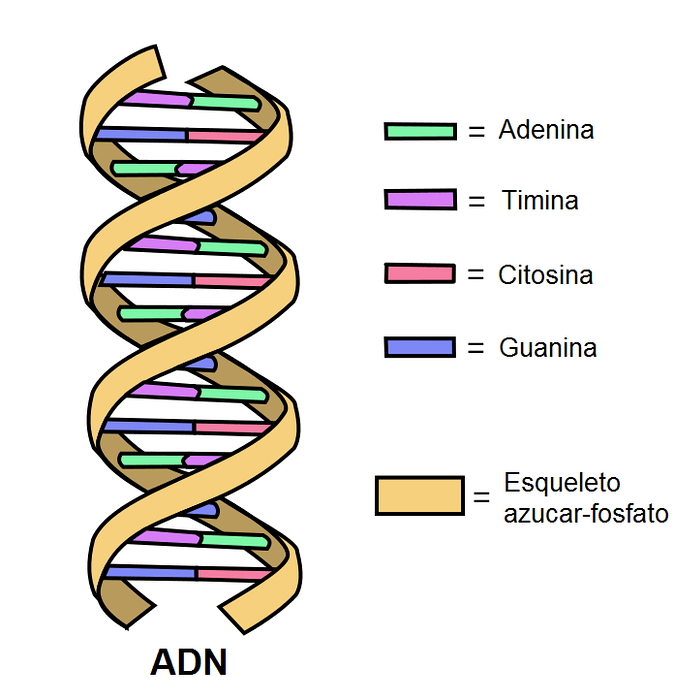
DNA består av to antiparallelle bånd som danner en dobbel helix. Hver av dem har et fosfodiesterbindingsskjelett som sukker knyttet til nitrogenholdige baser er festet på..
Inne i molekylet danner de nitrogenholdige basene i ett bånd hydrogenbindinger (to eller tre) med det komplementære båndet.
I et molekyl som dette viser de fleste viktige bindingsvinkler fri rotasjon. Nitrogenbase-sukker, sukker-fosfatgruppe og fosfodiesterbindinger er fleksible.
Dette gjør at DNA, sett på som en fleksibel stang, viser en viss evne til å bøye og vri. Denne fleksibiliteten tillater DNA å vedta komplekse lokale strukturer og danne interaksjonsløkker på korte, middels og lange avstander..
Denne fleksibiliteten forklarer også hvordan 2 meter DNA kan opprettholdes i hver diploide celle i et menneske. I en kjønnsceller (haploide celle) vil det være en meter DNA.
Selv om det ikke er en ubrytelig regel, eksisterer bakteriekromosomet som et enkelt superkveilet dobbeltbånd DNA-molekyl..
Den doble helixen vrir seg mer på seg selv (mer enn 10 bp per omdreining) og gir dermed en viss komprimering. Lokale knuter genereres også takket være manipulasjoner som er enzymatisk kontrollert.
I tillegg er det sekvenser i DNA som lar domener dannes i store sløyfer. Vi kaller strukturen som følge av superspolen og bestilte sløyfer for en nukleoid..
Disse gjennomgår dynamiske endringer takket være noen proteiner som gir noe strukturell stabilitet til det komprimerte kromosomet. Graden av komprimering i bakterier og archaea er så effektiv at det kan være mer enn ett kromosom per nukleoid.
Nukleoidet komprimerer det prokaryote DNA minst 1000 ganger. Den veldig topologiske strukturen til nukleoid er en grunnleggende del av reguleringen av genene som bæres av kromosomet. Det vil si at struktur og funksjon utgjør den samme enheten..
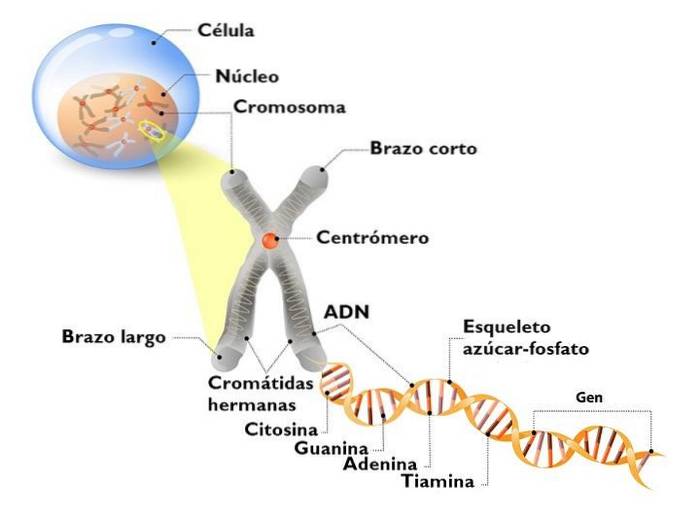
DNA i den eukaryote kjernen er ikke naken. Det samhandler med mange proteiner, hvorav de viktigste er histoner. Histoner er små, positivt ladede proteiner som binder seg til DNA ikke-spesifikt.
I kjernen er det vi observerer et komplekst DNA: histoner, som vi kaller kromatin. Det høyt kondenserte kromatinet, som vanligvis ikke uttrykkes, er heterokromatin. I motsetning er det minst komprimerte (løsere), eller eukromatin, kromatin med gener som uttrykkes.
Kromatin har forskjellige komprimeringsnivåer. Det mest elementære er nukleosom; Den følges av magnetfibre og interfase kromatinløkker. Det er først når et kromosom deler seg, at maksimale komprimeringsnivåer vises..
Nukleosomet er den grunnleggende organisasjonsenheten for kromatin. Hvert nukleosom består av en oktamer av histoner som danner en slags tromme..
Oktameren består av to kopier av hver av histonene H2A, H2B, H3 og H4. Rundt dem går DNA rundt 1,7 ganger. Det blir fulgt av en brøkdel av gratis DNA kalt en 20 bp linker assosiert med histon H1, og deretter et annet nukleosom. Mengden DNA i et nukleosom og det som binder det til et annet er omtrent 166 basepar.
Dette DNA-pakkingstrinnet komprimerer molekylet omtrent 7 ganger. Det vil si at vi går fra en meter til litt over 14 cm DNA.
Denne innpakningen er mulig fordi de positive histonene avbryter den negative ladningen av DNA, og den påfølgende elektrostatiske selvavstøtingen. Den andre årsaken er at DNA kan brettes på en slik måte at det kan snu oktameret i histoner..
Fiberen av perler i et halskjede dannet av mange suksessive nukleosomer er ytterligere viklet inn i en mer komprimert struktur.
Selv om vi ikke er klare på hvilken struktur den faktisk vedtar, vet vi at den når en tykkelse på ca. 30 nm. Dette er den såkalte 30 nm fiberen; Histon H1 er viktig for dannelsen og stabiliteten.
30 nm fiber er den grunnleggende strukturelle enheten til heterokromatin. Det av slappe nukleosomer, det for eukromatin.
30 nm fiberen er imidlertid ikke helt lineær. Tvert imot, det danner sløyfer på omtrent 300 nm i lengde, på en serpentin måte, på en lite kjent proteinmatrise..
Disse løkkene på en proteinmatrise danner en mer kompakt kromatinfiber med en diameter på 250 nm. Til slutt justeres de som en enkelt spiral på 700 nm tykk, noe som gir opphav til en av søsterkromatidene til et mitotisk kromosom..
Til slutt komprimerer DNA i kjernekromatin omtrent 10 000 ganger på kromosomet i delingscellen. I mellomfasekjernen er dens komprimering også høy, siden den er omtrent 1000 ganger sammenlignet med det "lineære" DNA..

I verden av utviklingsbiologi sies gametogenese å tilbakestille epigenomet. Det vil si at det sletter DNA-merkene som livet til personen som ga opphav til den gameten som ble produsert eller opplevd.
Disse kodene inkluderer DNA-metylering og kovalente modifikasjoner av histoner (Kode for histoner). Men ikke hele epigenomet tilbakestilles. Det som gjenstår med merker vil være ansvarlig for farens eller mors genetiske avtrykk.
Den implisitte tilbakestillingen til gametogenese er lettere å se i sædceller. I sæd er DNA ikke fullpakket med histoner. Derfor er informasjonen knyttet til endringene i produsentorganismen generelt ikke arvet..
I sæd er DNA pakket gjennom interaksjon med ikke-spesifikke DNA-bindende proteiner kalt protaminer. Disse proteinene danner disulfidbindinger med hverandre, og hjelper dermed til å danne overlappende DNA-lag som ikke frastøter elektrostatisk..



Ingen har kommentert denne artikkelen ennå.