De samvittighet eller kodeminant arv kan defineres som lik styrke mellom alleler. Hvis vi i ufullstendig dominans kan snakke om en genetisk doseringseffekt (AA>Aa>aa), i kodominans kan vi si at vi observerer den felles manifestasjonen av to produkter for samme karakter i samme individ, og med samme kraft.
En av grunnene som tillot Gregor Mendel å analysere på en enkel måte arvsmønstrene som ble observert av ham, var at tegnene som ble studert var fullstendig dominans.

Det er derfor i disse "klassiske" eller Mendel-tilfellene, genotypene AA Y Aa de manifesterer seg fenotypisk på samme måte (TIL dominerer helt til).
Men dette er ikke alltid tilfelle, og for monogene egenskaper (definert av et enkelt gen) kan vi finne to unntak som noen ganger kan forveksles: ufullstendig dominans og kodominans..
I den første, den heterozygote Aa manifesterer en fenotype som er mellomliggende i forhold til homozygoter AA Y aa; i det andre, som er det vi har å gjøre med her, manifesterer heterozygoten begge alleler, TIL Y til, med samme kraft, siden i virkeligheten ingen av dem er resessiv på den andre.
Artikkelindeks
Et av de beste eksemplene for å illustrere genetisk kodominans er blodgruppene i menneskelige populasjoner i henhold til ABO-klassifiseringssystemet..
I det praktiske liv blir en liten blodprøve utsatt for en responstest mot to antistoffer: anti-A-antistoffet og anti-B-antistoffet. A og B er navnene på to alternative former for det samme proteinet som er kodet på stedet Jeg; individer som ikke produserer noen form for proteinet, er homozygote recessive ii.
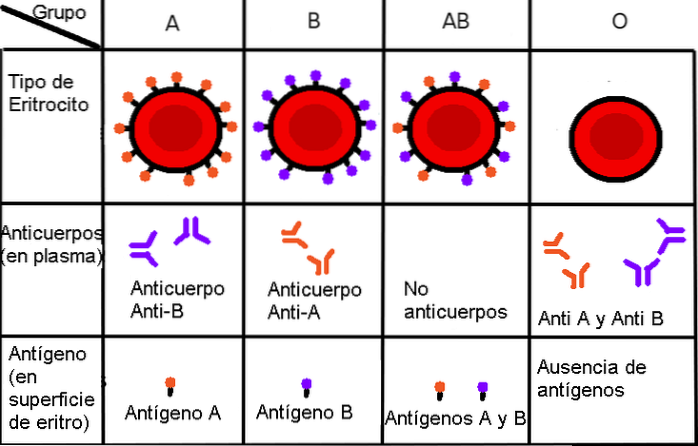
Derfor, ifølge ABO-systemet, er fenotypene til homozygote individer definert som følger:
1.- Personer hvis blod ikke gir noen immunrespons mot anti-A og anti-B antistoffer, er det fordi de ikke produserer protein A eller protein B, og derfor er de homozygote recessive ii.
Fenotypisk er dette individer med type O-blod, eller universelle givere, ettersom de ikke produserer noen av de to proteinene som kan forårsake immunavstøtning hos ikke-type O-blodmottakere. De fleste mennesker har denne typen blod..
2. - Tvert imot, hvis et individs blod reagerer med bare ett av antistoffene, Det er fordi den bare produserer en type av disse proteinene - det er derfor logisk sett at individet bare kan presentere to forskjellige genotyper.
Hvis det er et individ med type B-blod (og derfor ikke reagerer med anti-A-antistoffer, men bare med anti-B), kan hans genotype være homozygot JegBJegB, eller heterozygot JegBJeg (se neste avsnitt).
På samme måte kan individer som bare reagerer med anti-A-antistoffer være av genotypen JegTILJegTIL eller JegTILJeg. Så langt navigerer vi i kjente farvann, siden det er en type dominerende allel interaksjon i den reneste Mendelianske forstand: hvilken som helst allel Jeg (JegTIL eller JegB) vil dominere over allel i. Av denne grunn vil heterozygoter for A eller B være fenotypisk identiske med de som er homozygote for A eller B.
Heterozygoter for A og B, derimot, forteller oss en annen historie. Det vil si at et mindretall av den menneskelige befolkningen består av individer som reagerer med både anti-A og anti-B antistoffer; den eneste måten å vise denne fenotypen er ved å være genotypisk heterozygot JegTILJegB.
Derfor skapes et individ der ingen allel trekker seg tilbake ("forsvinner") og det er heller ikke "mellomliggende" mellom to andre: det er en ny fenotype, som vi kjenner som den universelle akseptoren, siden den ikke vil avvise noen type blod fra synspunkt til ABO-systemet.
For å fullføre forståelsen av kodominans, forstått som like styrke mellom alleler, er det nyttig å definere ufullstendig dominans. Den første tingen å avklare er at begge refererer til forhold mellom alleler av samme gen (og samme sted) og ikke til relasjoner eller geninteraksjoner mellom gener med forskjellige steder..
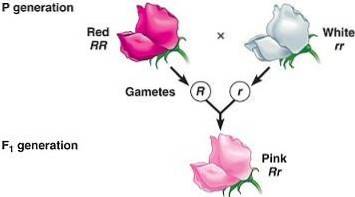
Den andre er at ufullstendig dominans manifesterer seg som et fenotypeprodukt av doseeffekten av produktet kodet av genet under analyse..
La oss ta et hypotetisk tilfelle av et monogent trekk der ett gen R, som koder for et monomert enzym, gir det opphav til en fargeforbindelse (eller pigment). Det recessive homozygote for det genet (rr), åpenbart, vil den mangle den fargen fordi den ikke gir opphav til enzymet som produserer det respektive pigmentet.
Både den dominerende homozygote RR som heterozygot Rr de vil manifestere farge, men på en annen måte: heterozygoten vil bli mer fortynnet siden den vil presentere halvparten av dosen av enzymet som er ansvarlig for å produsere pigmentet.
Det skal imidlertid forstås at genetisk analyse noen ganger er mer komplisert enn de enkle eksemplene som er gitt her, og at forskjellige forfattere tolker det samme fenomenet forskjellig..
Det er derfor mulig at i dihybride kryss (eller til og med med flere gener fra forskjellige lokus) kan de analyserte fenotypene vises i proporsjoner som ligner på et monohybridkors..
Bare streng og formell genetisk analyse kan tillate forskeren å konkludere hvor mange gener som er involvert i manifestasjonen av et trekk.
Historisk sett ble imidlertid begrepene kodominans og ufullstendig dominans brukt til å definere alleliske interaksjoner (gener fra samme sted), mens de som refererer til interaksjonene mellom gener fra forskjellige steder, eller geninteraksjoner per se, blir alle analysert som epistatiske interaksjoner.
Analysen av samspillet mellom forskjellige gener (av forskjellige lokus) som fører til manifestasjon av samme karakter kalles epistasisanalyse - det er i utgangspunktet ansvarlig for all genetisk analyse.



Ingen har kommentert denne artikkelen ennå.