EN fylogenetisk tre det er en grafisk matematisk fremstilling av historien og forfedrenes og etterkommernes forhold til grupper, populasjoner, arter eller andre taksonomiske kategorier. Teoretisk kan alle fylogenetiske trær grupperes i livets tre, og utgjør det universelle treet.
Disse grafiske fremstillingene har revolusjonert studiet av evolusjonærbiologi, siden de tillater etablering og definering av en art, testing av forskjellige evolusjonære hypoteser (som endosymbiotisk teori), evaluering av opprinnelsen til sykdommer (som HIV), etc..
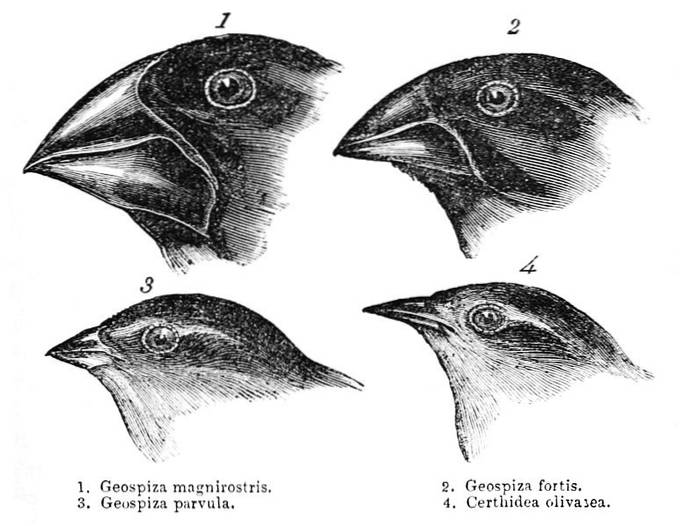
Trær kan rekonstrueres ved hjelp av morfologiske eller molekylære tegn, eller begge deler. På samme måte er det forskjellige metoder for å bygge dem, den vanligste er den kladistiske metodikken. Dette søker å identifisere delte avledede tegn, kjent som synapomorphy.
Artikkelindeks
Et av prinsippene utviklet av Charles Darwin utgjør det felles forfedre til alle levende organismer - det vil si at vi alle deler en fjern forfedre.
På "Opprinnelsen til arter" Darwin løfter metaforen til "livets tre." Faktisk bruker han et hypotetisk grafisk tre for å utvikle ideen sin (merkelig nok er det den eneste illustrasjonen av Kilde).
Representasjonen av denne metaforen er det vi kjenner som fylogenetiske trær, som lar oss grafisk vise historien og forholdet til en bestemt gruppe organismer..
I fylogenetiske trær kan vi skille mellom følgende deler - fortsetter med den botaniske analogien:
Grener: Linjene på treet kalles "grener", og disse representerer studiepopulasjonene over tid. Avhengig av tretype (se nedenfor), kan grenens lengde eller ikke ha noen betydning.
På tipsene til grenene finner vi organismer som vi vil evaluere. Dette kan være enheter som for tiden er i live, eller utdødde vesener. Arten vil være bladene på treet vårt.
Rot: roten er den eldste grenen av treet. Noen har det og kalles rotte trær, mens andre ikke har det.
Noder: grenpunktene til grenene i to eller flere linjer kalles noder. Prikken representerer den siste felles forfedren til etterkommergruppene (merk at disse forfedrene er hypotetiske).
Eksistensen av en node innebærer en spesifikasjonshendelse - opprettelse av nye arter. Etter dette følger hver art sin evolusjonære forløp.
I tillegg til disse tre grunnleggende begrepene, er det andre nødvendige begreper når det gjelder fylogenetiske trær:
Politomi: når et fylogenetisk tre har mer enn to grener i en node, sies det at det er en polytomi. I disse tilfellene er det fylogenetiske treet ikke fullstendig løst, fordi forholdet mellom de involverte organismer ikke er klart. Dette skyldes vanligvis mangel på data, og kan bare løses når en forsker akkumulerer mer.
Ekstern gruppe: i fylogenetiske emner er det vanlig å høre begrepet ekstern gruppe - også kalt utgruppe. Denne gruppen er valgt for å kunne rote treet. Det bør velges som en takson som tidligere avvek fra studiegruppen. For eksempel, hvis jeg studerer pigghuder, kan du bruke utgruppe sjøspruter.
Det er tre grunnleggende typer trær: kladogrammer, additivtrær og ultrametriske trær..
Kladogrammer er de enkleste trærne og viser forholdet til organismer når det gjelder felles forfedre. Informasjonen til denne typen trær ligger i forgreningsmønstrene, siden størrelsen på grenene ikke har noen ekstra betydning.
Den andre tretypen er tilsetningsstoffet, også kalt metriske trær eller fylogrammer. Lengden på grenene er relatert til mengden evolusjonær endring.
Til slutt har vi ultrametriske trær eller dendogrammer, der alle tuppen av trærne er på samme avstand (noe som ikke er tilfelle i fylogrammet, der et tips kan vises lavere eller høyere enn partneren). Grenens lengde er relatert til evolusjonstid.
Valget av treet er direkte knyttet til evolusjonsspørsmålet som vi ønsker å svare på. For eksempel, hvis vi bare bryr oss om forholdet mellom individer, vil et kladogram være tilstrekkelig for studien.
Selv om fylogenetiske trær ofte er mye brukt grafer i evolusjonær biologi (og generell biologi), er det mange studenter og utøvere som feiltolker budskapet om at disse tilsynelatende enkle grafene er ment å levere til leseren..
Den første feilen er å lese dem sidelengs, forutsatt at evolusjon innebærer fremgang. Hvis vi forstår evolusjonsprosessen riktig, er det ingen grunn til å tro at de forfedre artene er til venstre og de mer avanserte artene til høyre..
Selv om den botaniske analogien til et tre er veldig nyttig, kommer det et punkt der det ikke lenger er så nøyaktig. Det er en avgjørende trestruktur som ikke er tilstede i treet: stammen. I fylogenetiske trær finner vi ingen hovedgrener.
Spesielt kan noen mennesker betrakte mennesket som det endelige "målet" for evolusjonen, og derfor arten. Homo sapiens bør alltid være lokalisert.
Dette synet er imidlertid ikke i samsvar med evolusjonære prinsipper. Hvis vi forstår at fylogenetiske trær er mobile elementer, kan vi plassere Homo i hvilken som helst terminal posisjon på treet, siden denne karakteristikken ikke er relevant i representasjonen.
En viktig egenskap som vi må forstå om fylogenetiske trær er at de representerer ikke-statiske grafer..
I dem kan alle disse grenene rotere - på samme måte som en mobil kan gjøre det. Vi mener ikke at vi kan flytte grenene etter eget ønske, fordi noen bevegelser vil innebære endring av mønsteret eller topologi Fra treet. Det vi kan rotere er nodene.
For å tolke budskapet til et tre, må vi ikke fokusere på grenene, vi må fokusere på grenpunktene, som er det viktigste aspektet av grafen.
I tillegg må vi huske på at det er flere måter å tegne et tre på. Mange ganger avhenger det av stilen på boken eller magasinet, og endringer i formen og posisjonen til grenene påvirker ikke informasjonen de vil formidle til oss..
Når skal vi referere til arter nåværende vi skal ikke bruke forfedres konnotasjoner til dem. For eksempel, når vi tenker på forholdet mellom sjimpanser og mennesker, kunne vi feilaktig forstå at sjimpanser er forfedre med hensyn til vår slekt..
Imidlertid var den vanlige forfederen til sjimpanser og mennesker ingen av dem. Å tro at sjimpansen er forfedret, ville være å anta at utviklingen stoppet når begge linjene ble skilt..
Etter den samme logikken til disse ideene, forteller et fylogenetisk tre oss ikke om det er unge arter heller. Ettersom de alleliske frekvensene er i konstant endring og det er nye tegn som endrer seg over tid, er det vanskelig å bestemme alderen til en art, og absolutt, et tre gir oss ikke slike data.
"Endringen i allelfrekvenser over tid" er hvordan populasjonsgenetikk definerer evolusjon.
Når vi ser på et fylogenetisk tre, må vi forstå at denne grafen ganske enkelt er en hypotese generert fra konkrete bevis. Det kan være at hvis vi legger til flere tegn i treet, endrer det topologien.
Vitenskapenes ekspertise i å velge de beste karakterene for å belyse forholdene til de aktuelle organismer er nøkkelen. I tillegg er det veldig kraftige statistiske verktøy som gjør det mulig for forskere å evaluere trær og velge det mest sannsynlige..
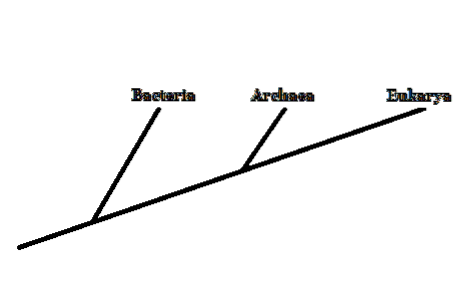
I 1977 foreslo forskeren Carl Woese å gruppere levende organismer i tre domener: Archaea, Bacteria og Eukarya. Dette nye klassifiseringssystemet (tidligere var det bare to kategorier, Eukaryota og Prokaryota) var basert på den ribosomale RNA-molekylære markøren.
Bakterier og eukaryoter er allment kjente organismer. Archaea forveksles ofte med bakterier. Imidlertid er disse veldig forskjellige i strukturen til deres mobilkomponenter..
Derfor, selv om de er mikroskopiske organismer som bakterier, er medlemmer av Archaea-domenet nærmere knyttet til eukaryoter - fordi de deler en nærmere felles forfader..
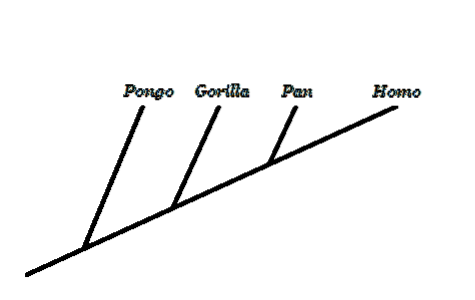
Innen evolusjonær biologi er et av de mest kontroversielle temaene menneskets evolusjon. For motstanderne av denne teorien er ikke en evolusjon med utgangspunkt i en apelike forfader som ga opphav til det moderne mennesket, logisk..
Et sentralt begrep er å forstå at vi ikke utviklet oss fra nåværende aper, men vi deler en felles forfader med dem. I apenes og menneskets tre skiller det seg ut at det vi kjenner som "ape" ikke er en gyldig monofyletisk gruppe, siden det ekskluderer mennesker.
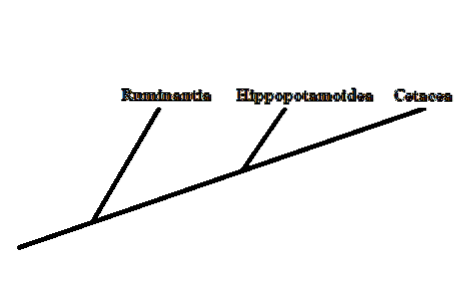
Evolusjonelt sett representerte hvaler en gruppe virveldyr hvis forhold til resten av deres andre pattedyr ikke var veldig tydelige. Morfologisk har hvaler, delfiner og andre medlemmer få likheter med resten av pattedyrene.
For øyeblikket, takket være studiet av forskjellige morfologiske og molekylære tegn, har det vært mulig å konkludere med at søstergruppen til de store hvaler ble dannet av artiodaktylene - hovdyr med jevne hov..



Ingen har kommentert denne artikkelen ennå.