
De genetisk rekombinasjon det er prosessen der nukleinsyremolekyler utveksler fragmenter og genererer et nytt molekyl. Det er veldig vanlig i DNA, men RNA er også et substrat for rekombinasjon. Rekombinasjon er, etter mutasjon, den viktigste kilden til generering av genetisk variasjon.
DNA deltar i forskjellige biokjemiske prosesser. Under replikasjon fungerer den som en mal for generering av to nye DNA-molekyler. I transkripsjon tillater det generering av RNA-molekyler fra spesifikke regioner kontrollert av en promoter.
Men i tillegg til det, er DNA også i stand til å utveksle fragmenter. Gjennom denne prosessen genererer den nye kombinasjoner som ikke er et produkt av de to foregående prosessene, og heller ikke av befruktning..
All rekombinasjonsprosess innebærer å bryte og sammenføye DNA-molekylene som deltar i prosessen. Denne mekanismen varierer avhengig av rekombinasjonssubstratet, enzymene som deltar i prosessen og mekanismen for utførelse..
Rekombinasjon avhenger vanligvis av eksistensen av komplementære, like (om ikke identiske) eller homologe regioner mellom de rekombinerende molekylene. I tilfelle molekyler rekombineres i prosesser som ikke styres av homologi, sies rekombinasjonen å være ikke-homolog.
Hvis homologien innebærer en veldig kort region som er tilstede i begge molekylene, sies rekombinasjonen å være stedsspesifikk..
Artikkelindeks
Det vi kaller homologi i rekombinasjon, refererer ikke nødvendigvis til evolusjonær opprinnelse til de deltakende molekylene. Vi snakker mer om graden av likhet i nukleotidsekvensen.
Ikke-reparativ rekombinasjon forekommer for eksempel i eukaryoter under meiose-prosessen. Utvilsomt kan det ikke være større homologi enn mellom par av kromosomer i samme celle.
Det er derfor de kalles homologe kromosomer. Imidlertid er det tilfeller der DNA av en celle bytter materiale med et fremmed DNA. Disse DNAene må være veldig like rekombinerte, men de trenger ikke nødvendigvis å dele samme forfedre (homologi) for å oppnå det..
Feste- og utvekslingsstedet mellom to DNA-molekyler kalles chiasme, og selve prosessen kalles tverrbinding. Under overgangen verifiseres en utveksling av bånd mellom de deltakende DNA-ene..
Dette genererer et mynt integrert, som er to DNA-molekyler som fysisk er bundet til ett. Når man "løser" mynt integreres, genereres to molekyler, generelt endret (rekombinant).
Å "løse", i sammenheng med rekombinasjon, er å skille komponent-DNA-molekylene til et mynt integrert.
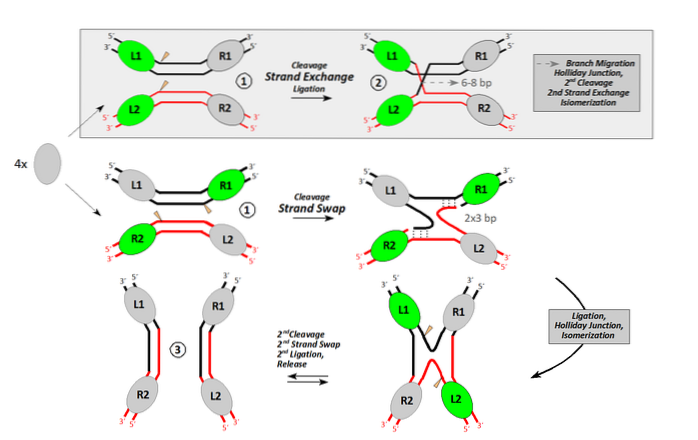
I stedsspesifikk rekombinasjon har to DNA-molekyler, vanligvis ikke homologe, en kort sekvens som er felles for begge. Denne sekvensen er målet for et spesifikt spleisningsenzym.
Enzymet, som er i stand til å gjenkjenne denne sekvensen og ikke en annen, kutter det på et bestemt sted i begge molekylene. Ved hjelp av noen andre faktorer utveksler den DNA-båndene til de to deltakende molekylene og danner et mynt integrert.
Dette er grunnlaget for dannelsen av myntet integreres mellom genomet til bakterien Escherichia coli og den fra bakteriofagen lambda. En bakteriofag er et virus som infiserer bakterier.
Dannelsen av dette myntintegratet utføres av et enzym kodet i virusgenomet: lambda integrase. Den gjenkjenner en vanlig sekvens som kalles attP i det sirkulære genomet til viruset, og attB i bakterienes.
Ved å kutte begge sekvensene i begge molekylene, genererer det frie segmenter, bytter bånd og blir med de to genomene. En større eller myntintegrert sirkel dannes.
I myntintegrasjon bæres virusgenomet passivt av bakteriegenomet, som det replikerer med. I denne tilstanden sies det at viruset er i provirus-tilstand, og at bakterien er lysogen for den..
Den omvendte prosessen, det vil si oppløsningen til myntintegreringen, kan ta mange generasjoner - eller til og med ikke forekomme. Imidlertid medieres dette enzymatisk av et annet protein kodet av virusgenomet kalt excisionase. Når dette skjer, skiller viruset seg fra mynt integreres, reaktiverer og forårsaker cellelyse..
Homolog rekombinasjon skjer mellom DNA-molekyler som deler minst omtrent 40 nukleotider med fullstendig eller nesten fullstendig likhet. For å gjennomføre rekombinasjonsprosessen, må minst én endonuklease delta.
Endonukleaser er enzymer som lager interne kutt i DNA. Noen gjør det for å fortsette med å nedbryte DNA. Andre, som i tilfelle rekombinasjon, gjør det for å generere en bulke i DNA.
Dette unike nicket tillater behandling av et enkeltbånds DNA med en fri ende. Denne frie enden, orientert av en rekombinase, gjør det mulig for et enkelt bånd å invadere et dobbelt DNA, og fortrenger det bånd som er identisk med det..
Dette er krysspunktet mellom et donor ("inntrenger") DNA-molekyl og en annen reseptor..
Enzymet (rekombinase) som utfører prosessen med båndinvasjon og utveksling Escherichia coli det kalles RecA. Det er andre homologe proteiner i prokaryoter, slik som RadA i archaea. I eukaryoter kalles det tilsvarende enzymet RAD51.
Når det invasive båndet fortrenger beboeren, samhandler hun med båndet som var igjen i donormolekylet. Begge steder forsegles ved virkning av en ligase.
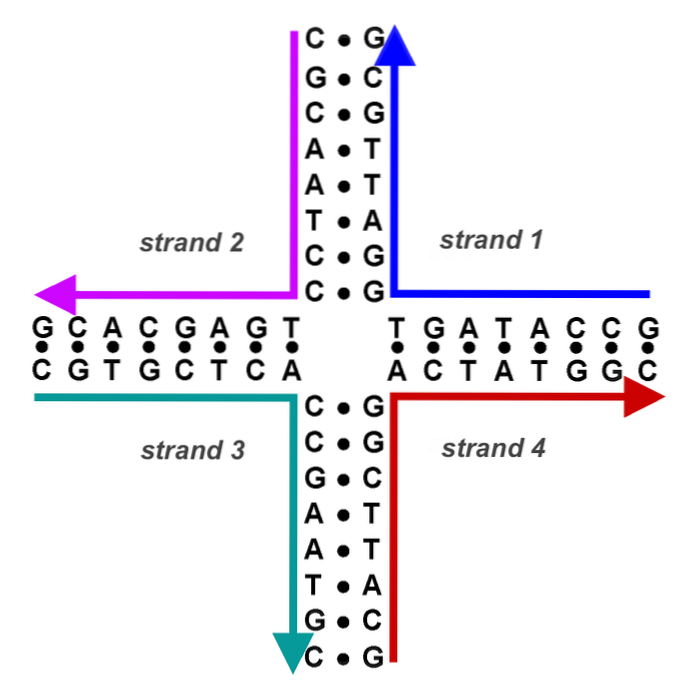
Nå har vi et DNA av hybridbånd (et giverbånd og et mottakerbånd, av forskjellig opprinnelse) flankert av donor-DNA og mottaker-DNA. Crossover-punktene (chiasmata) beveger seg i begge retninger med minst 200 bp.
Hvert punkt med kryssdannelse danner det som er kjent som Holliday-strukturen (korsformet DNA-produkt fra en rekombinasjonshendelse).
Dette korsformede DNA må løses av andre endonukleaser. Chimerisk eller hybrid DNA med denne strukturen kan løses på to måter. Hvis det andre endonukleotidsnittet skjer i samme bånd som det første, forekommer ingen rekombinasjon. Hvis det andre kuttet skjer i det andre båndet, er de resulterende produktene rekombinante.
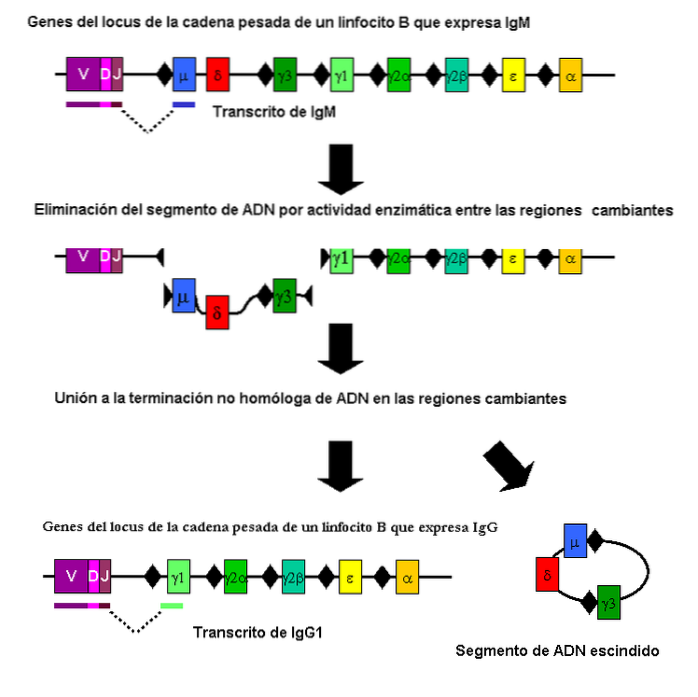
Dette er en type somatisk rekombinasjon (ikke meiotisk) som bidrar til generasjonen i den enorme variasjonen av antistoffer mot immunsystemet.
Denne rekombinasjonen finner sted i spesielle fragmenter av genene som koder for polypeptidkjedene som definerer dem. Den utføres av B-celler og involverer forskjellige genetiske regioner.
Interessant, det er parasitter som Trypanosoma brucei som benytter en lignende rekombinasjonsmekanisme for å skape variasjon i et overflateantigen. På denne måten kan de unngå vertsresponsen hvis verten ikke klarer å generere antistoffet som er i stand til å gjenkjenne det "nye" antigenet..
Til slutt er det rekombinasjonsprosesser som ikke avhenger av likheten i sekvensen til de deltakende molekylene. I eukaryoter er for eksempel rekombinasjon av ikke-homologe ender veldig viktig.
Dette skjer med DNA-fragmenter som viser dobbeltbåndsbrudd i DNA. Disse blir "reparert" ved at cellen forbinder dem med andre fragmenter også med dobbeltbåndsbrudd.
Imidlertid trenger disse molekylene ikke nødvendigvis å være like for å delta i denne rekombinasjonsprosessen. Det vil si at ved å reparere skaden kan cellen slutte seg til ikke-relaterte DNA, og dermed skape et virkelig nytt (rekombinant) molekyl.
Rekombinasjon sikrer troskapen til DNA-informasjon under og etter replikasjonsprosessen. Rekombinasjon oppdager DNA-skade under den nye båndprosessen i dette ekstremt lange makromolekylet.
Siden hvert band har sin egen informasjon og komplementær, garanterer rekombinasjon at ingen går tapt. Hver fungerer som et vitne for den andre. På samme måte, i diploide organismer, er et homologt kromosom vitne til søsken, og omvendt.
På den annen side, når DNA har blitt replikert, er skadereparasjonsmekanismene til dette molekylet variert. Noen er direkte (skaden blir direkte handlet på) og andre er indirekte.
Indirekte reparasjonsmekanismer er avhengig av at rekombinasjon skal finne sted. Det vil si at for å reparere skaden i et DNA-molekyl, brukes et annet homologt molekyl. Dette vil fungere i reparativ rekombinasjon som en mal som den har blitt skadet av..
Rekombinasjon er i stand til å skape enorm kromosomal variasjon under meiose. Somatisk rekombinasjon genererer også variasjon, som i tilfelle antistoffer hos virveldyr.
I mange organismer er meiose gametisk. I seksuelt reproduserte organismer er rekombinasjon en av de kraftigste måtene å generere variabilitet på.
Det vil si at til spontan mutasjon og kromosomsegregering, må rekombinasjon legges til som et annet element som genererer gametisk variasjon..
Integreringen av bakteriofaggenomer ved stedsspesifikk rekombinasjon har derimot bidratt til ombygging av genomet til deres vertsbakterier..
Dette har bidratt til generasjonen av genomisk variasjon og evolusjon av denne viktige gruppen av levende vesener..
Vi har allerede sett at DNA kan repareres, men ikke hva som skader det. I virkeligheten kan nesten alt skade DNA, startende med feil replikering som blir ukorrigert..
Men utover det kan DNA bli skadet av UV-lys, ioniserende stråling, frie oksygenradikaler fra cellulær respirasjon, og det vi spiser, røyker, puster, inntar eller berører..
Heldigvis trenger du ikke å gi opp å leve for å beskytte DNA. Visse ting må oppgis, men det store arbeidet gjøres av selve cellen. Disse mekanismene for å oppdage skade på DNA og reparere det, har åpenbart et genetisk grunnlag, og deres mangel, enorme konsekvenser.
Sykdommer relatert til defekter i homolog rekombinasjon inkluderer for eksempel Bloom- og Werner-syndromer, familiær bryst- og eggstokkreft, etc..


Ingen har kommentert denne artikkelen ennå.