De ribose Det er et sukker med fem karbon som er tilstede i ribonukleosider, ribonukleotider og deres derivater. Det kan bli funnet med andre navn som β-D-ribofuranose, D-ribose og L-ribose.
Nukleotider er "byggesteinene" i ribonukleinsyre (RNA) ryggraden. Hvert nukleotid består av en base som kan være adenin, guanin, cytosin eller uracil, en fosfatgruppe og et sukker, ribose.

Denne typen sukker er spesielt rikelig i muskelvev, der den er assosiert med ribonukleotider, spesielt med adenosintrifosfat eller ATP, som er viktig for muskelfunksjon..
D-ribose ble oppdaget i 1891 av Emil Fischer, og siden den gang har mye oppmerksomhet blitt lagt på dens fysisk-kjemiske egenskaper og dens rolle i cellulær metabolisme, det vil si som en del av skjelettet av ribonukleinsyre, ATP og forskjellige koenzymer.
Først ble dette bare oppnådd fra hydrolyse av gjær-RNA, til det på 1950-tallet klarte å bli syntetisert fra D-glukose i mer eller mindre tilgjengelige mengder, noe som tillot industrialisering av produksjonen..
Artikkelindeks
Ribose er en aldopentose som ofte ekstraheres som en ren kjemisk forbindelse i form av D-ribose. Det er et organisk stoff løselig i vann, med et hvitt og krystallinsk utseende. Å være et karbohydrat, har ribose polære og hydrofile egenskaper.
Ribose oppfyller den vanlige karbohydratregelen: den har samme antall karbon og oksygenatomer, og dobbelt så mange i hydrogenatomer..
Gjennom karbonatomene i posisjon 3 eller 5 kan dette sukkeret binde seg til en fosfatgruppe, og hvis det binder seg til en av de nitrogenholdige basene av RNA, dannes et nukleotid.
Den vanligste måten å finne ribose i naturen er som D-ribose og 2-deoksy-D-ribose, disse er komponenter av nukleotider og nukleinsyrer. D-ribose er en del av ribonukleinsyre (RNA) og 2-deoksy-D-ribose av deoksyribonukleinsyre (DNA).
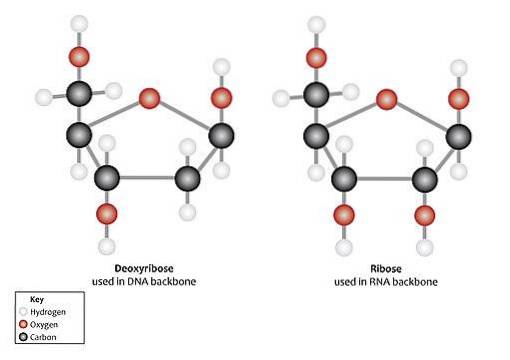
Ved nukleotider er begge typer pentose i β-furanose-form (lukket femkantet ring).
I løsning er fri ribose i likevekt mellom aldehydformen (åpen kjede) og den sykliske β-furanoseformen. Imidlertid inneholder RNA bare den sykliske formen β-D-ribofuranose. Den biologisk aktive formen er vanligvis D-ribose.
Ribose er et sukker avledet av glukose som tilhører gruppen aldopentoser. Molekylformelen er C5H10O5 og har en molekylvekt på 150,13 g / mol. Siden det er et monosakkaridsukker, skiller dens hydrolyse molekylet i dets funksjonelle grupper.
Den har, som formelen indikerer, fem karbonatomer som kan bli funnet syklisk som en del av fem- eller seksleddet ringer. Dette sukkeret har en aldehydgruppe ved karbon 1 og en hydroksylgruppe (-OH) ved karbonatomer fra posisjon 2 til posisjon 5 i pentosringen.
Ribosemolekylet kan representeres i Fisher-projeksjonen på to måter: D-ribose eller L-ribose, med form L som stereoisomer og enantiomer av form D og omvendt..
Klassifiseringen av D- eller L-formen avhenger av orienteringen av hydroksylgruppene i det første karbonatomet etter aldehydgruppen. Hvis denne gruppen er orientert mot høyre side, tilsvarer molekylet i Fishers representasjon D-ribose, ellers hvis det er mot venstre side (L-ribose).
Haworth-projeksjonen av ribose kan representeres i to ytterligere strukturer avhengig av orienteringen av hydroksylgruppen på karbonatomet som er anomer. I β-stillingen er hydroksylen orientert mot toppen av molekylet, mens α-posisjonen orienterer hydroksylen mot bunnen.

I følge Haworth-projeksjonen kan det således være fire mulige former: β-D-ribose, α-D-ribose, β-L-ribose eller α-L-ribose.
Når fosfatgrupper er festet til ribose, blir disse ofte referert til som α, β og Ƴ. Hydrolyse av nukleosidtrifosfat gir den kjemiske energien til å drive et bredt utvalg av cellulære reaksjoner.
Det er blitt foreslått at ribosefosfat, produkt fra nedbrytning av ribonukleotider, er en av hovedforløperne til furan og tiofenoler, som er ansvarlig for den karakteristiske lukten av kjøtt..
Den kjemiske plastisiteten til ribose gjør at molekylet er involvert i de aller fleste biokjemiske prosesser inne i cellen, noen som oversettelse av DNA, syntese av aminosyrer og nukleotider, etc..
Ribose fungerer kontinuerlig som et kjemisk middel inne i cellen, siden nukleotider kan ha en, to eller tre fosfatgrupper kovalent bundet til hverandre ved vannfrie bindinger. Disse er kjent som henholdsvis nukleosider mono-, di- og trifosfat..
Bindingen mellom ribose og fosfat er av estertypen, hydrolysen av denne bindingen frigjør omtrent 14 kJ / mol under standardbetingelser, mens den for hver av anhydridbindinger frigjør omtrent 30 kJ / mol.
I ribosomer, for eksempel, kan 2'-hydroksylgruppen av ribose danne en hydrogenbinding med de forskjellige aminosyrene, en forening som tillater syntese av proteiner fra tRNA i alle kjente levende organismer.
Giftet til de fleste slanger inneholder en fosfodiesterase som hydrolyserer nukleotider fra 3'-enden som har fri hydroksyl, og bryter bindingen mellom 3'-hydroksylen av ribose eller deoksyribose.
I medisinske sammenhenger brukes det til å forbedre ytelse og treningskapasitet ved å øke muskelenergi. Kronisk utmattelsessyndrom behandles også med dette sakkaridet, samt fibromyalgi og visse kranspulsårssykdommer..
Forebyggende brukes den til å forhindre muskeltretthet, kramper, smerte og stivhet etter trening hos pasienter med arvelig lidelse av myoadenylatdeaminase-mangel eller AMP-deaminase-mangel..



Ingen har kommentert denne artikkelen ennå.