
De DNA-oversettelse er prosessen der informasjonen i messenger-RNAene produsert under transkripsjon (kopien av informasjonen i en DNA-sekvens i form av RNA) blir "oversatt" til en aminosyresekvens ved hjelp av proteinsyntese.
Fra et cellulært perspektiv er genuttrykk en relativt kompleks affære som skjer i to trinn: transkripsjon og oversettelse..
Alle gener som blir uttrykt (uansett om de koder for peptidsekvenser, det vil si proteiner) gjør det i utgangspunktet ved å overføre informasjonen i DNA-sekvensen til et messenger RNA (mRNA) -molekyl gjennom en prosess som kalles transkripsjon.
Transkripsjon oppnås ved spesielle enzymer kjent som RNA-polymeraser, som bruker en av de komplementære strengene i genets DNA som en mal for syntesen av et "pre-mRNA" -molekyl, som deretter behandles for å danne et modent mRNA..
For gener som koder for proteiner, blir informasjonen i modne mRNAs "lest" og oversatt til aminosyrer i henhold til den genetiske koden, som spesifiserer hvilket kodon eller nukleotidtriplett som tilsvarer hvilken bestemt aminosyre..
Spesifikasjonen av aminosyresekvensen til et protein, avhenger derfor av den innledende sekvensen av nitrogenholdige baser i DNA som tilsvarer genet og deretter i mRNA som bærer denne informasjonen fra kjernen til cytosolen (i eukaryote celler); prosess som også er definert som mRNA-styrt proteinsyntese.
Gitt at det er 64 mulige kombinasjoner av de 4 nitrogenholdige basene som utgjør DNA og RNA og bare 20 aminosyrer, kan en aminosyre kodes av forskjellige tripletter (kodoner), og det er derfor den genetiske koden sies å være "degenerert" (unntatt aminosyren metionin, som er kodet av et unikt AUG-kodon).
Artikkelindeks
I eukaryote celler foregår transkripsjon i kjernen og translasjon i cytosolen, slik at mRNAene som dannes under den første prosessen også spiller en rolle i transporten av informasjon fra kjernen til cytosolen, der cellene finnes. Biosyntetisk maskineri (ribosomer).
Det er viktig å nevne at avdelingen av transkripsjon og oversettelse i eukaryoter er sant for kjernen, men det er ikke det samme for organeller med sitt eget genom som kloroplaster og mitokondrier, som har systemer som ligner mer på de av prokaryote organismer..
Eukaryote celler har også cytosoliske ribosomer festet til membranene i det endoplasmatiske retikulum (grovt endoplasmatisk retikulum), der oversettelsen av proteiner som er bestemt for å sette inn i cellemembraner eller som krever posttranslasjonsbehandling som forekommer i nevnte rom, forekommer..
MRNAene modifiseres i endene når de transkriberes:
- Når 5'-enden av mRNA kommer ut fra overflaten av RNA-polymerase II under transkripsjon, blir den umiddelbart "angrepet" av en gruppe enzymer som syntetiserer en "hette" sammensatt av 7-metylguanylat og som er koblet til det terminale nukleotidet av mRNA via en 5 ', 5' trifosfatbinding.
- 3'-enden av mRNA gjennomgår en "spalting" av en endonuklease, som genererer en 3'-fri hydroksylgruppe som en "streng" eller "hale" av adeninrester (fra 100 til 250) er festet til, som tilsettes en samtidig av et enzym poly (A) polymerase.
"Hetten 5 '" og "halen poly A ”utfører funksjoner i beskyttelsen av mRNA-molekyler mot nedbrytning, og i tillegg fungerer de i transport av modne transkripsjoner mot henholdsvis cytosolen og i initiering og avslutning av translasjon..
Etter transkripsjon gjennomgår de "primære" mRNAene med sine to modifiserte ender, fremdeles til stede i kjernen, en "spleising" -prosess hvor introniske sekvenser vanligvis fjernes og de resulterende eksonene blir sammenføyd (post-transkripsjonsbehandling), med hvilke modne transkripsjoner blir oppnådd som forlater kjernen og når cytosolen.
Skjøting utføres av et riboproteinkompleks kalt spleisosom (Anglisisme av spleisosom), bestående av fem små ribonukleoproteiner og RNA-molekyler, som er i stand til å "gjenkjenne" regionene som skal fjernes fra det primære transkriptet.
I mange eukaryoter er det et fenomen kjent som "alternativ spleising", noe som betyr at forskjellige typer posttranskripsjonelle modifikasjoner kan produsere forskjellige proteiner eller isoenzymer som skiller seg fra hverandre i noen aspekter av deres sekvenser..
Når modne transkripsjoner forlater kjernen og transporteres for translasjon i cytosolen, blir de behandlet av det translasjonelle komplekset kjent som ribosomet, som består av et kompleks av proteiner assosiert med RNA-molekyler..
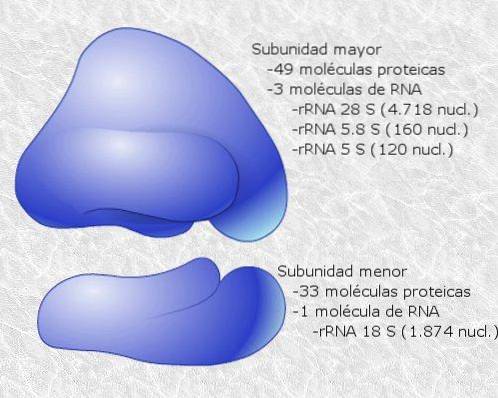
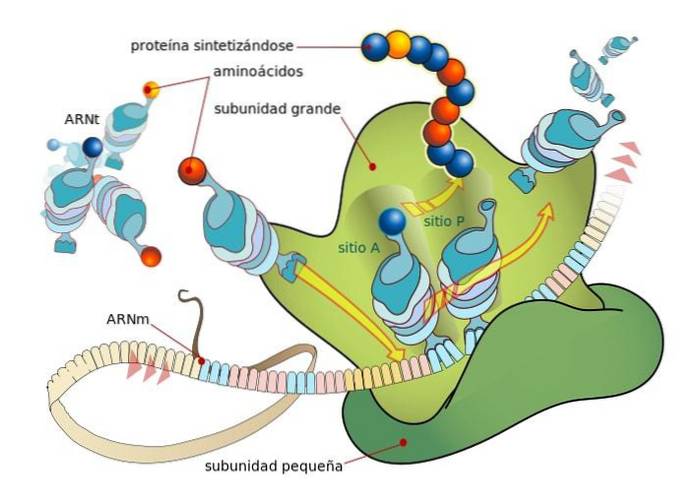
Ribosomer er sammensatt av to underenheter, den ene "stor" og den andre "liten", som er fritt dissosiert i cytosolen og binder eller assosieres på mRNA-molekylet som oversettes.
Bindingen mellom ribosomer og mRNA avhenger av spesialiserte RNA-molekyler som assosieres med ribosomale proteiner (ribosomalt RNA eller rRNA og overfører RNA eller tRNA), som hver har spesifikke funksjoner..
TRNA er molekylære "adaptere", siden de gjennom en av endene kan "lese" hvert kodon eller triplett i modent mRNA (ved basekomplementaritet) og gjennom den andre kan de binde seg til aminosyren som er kodet av "lest" kodonet..
RRNA-molekylene er derimot ansvarlige for å akselerere (katalysere) bindingsprosessen til hver aminosyre i den begynnende peptidkjeden.
Et modent eukaryotisk mRNA kan "leses" av mange ribosomer, så mange ganger som cellen indikerer. Med andre ord kan det samme mRNA gi mange kopier av det samme proteinet..
Når et modent mRNA nærmer seg av ribosomale underenheter, "skanner" riboproteinkomplekset sekvensen til nevnte molekyl til den finner et startkodon, som alltid er AUG og involverer innføring av en metioninrest.
AUG-kodonen definerer leserammen for hvert gen og definerer i tillegg den første aminosyren til alle proteiner som er oversatt i naturen (denne aminosyren blir ofte eliminert etter translasjonelt).
Tre andre kodoner er identifisert som de som induserer oversettelsesterminering: UAA, UAG og UGA..
De mutasjonene som involverer en endring av nitrogenholdige baser i tripletten som koder for en aminosyre og som resulterer i stoppkodoner, er kjent som tullmutasjoner, siden de forårsaker en tidlig arrestasjon av synteseprosessen, som danner kortere proteiner..
Nær 5'-enden av modne mRNA-molekyler er det regioner som ikke er oversatt (UTR). Ikke oversatt region), også kalt "leder" -sekvenser, som ligger mellom det første nukleotidet og oversettelsesstartkodonet (AUG).
Disse UTR-områdene som ikke er oversatt, har spesifikke bindingssteder med ribosomer og hos mennesker er for eksempel omtrent 170 nukleotider i lengde, blant hvilke det er reguleringsregioner, proteinbindingsseter som fungerer i reguleringen av oversettelsen, osv..
Oversettelse, så vel som transkripsjon, består av 3 faser: en initieringsfase, en forlengelsesfase og til slutt en avslutningsfase..
Den består av montering av translasjonskomplekset på mRNA, som fortjener foreningen av tre proteiner kjent som initieringsfaktorer (IF). Initiasjonsfaktor) IF1, IF2 og IF3 til den lille underenheten til ribosomet.
"Pre-initiering" -komplekset dannet av initieringsfaktorene og den lille ribosomale underenheten, binder i sin tur med et tRNA som "bærer" en metioninrest, og dette molekylsettet binder seg til mRNA, nær startkodonet. AUG.
Disse hendelsene fører til binding av mRNA til den store ribosomale underenheten, noe som fører til frigjøring av initieringsfaktorer. Den store ribosomsubenheten har 3 bindingssteder for tRNA-molekyler: A-setet (aminosyre), P-stedet (polypeptid) og E-stedet (utgang)..
Sted A binder seg til antikodonet til aminoacyl-tRNA som er komplementært til det for mRNA som blir oversatt; P-stedet er der aminosyren overføres fra tRNA til det begynnende peptidet, og E-stedet er der den finnes i "tomt" tRNA før den frigjøres i cytosolen etter at aminosyren er levert.
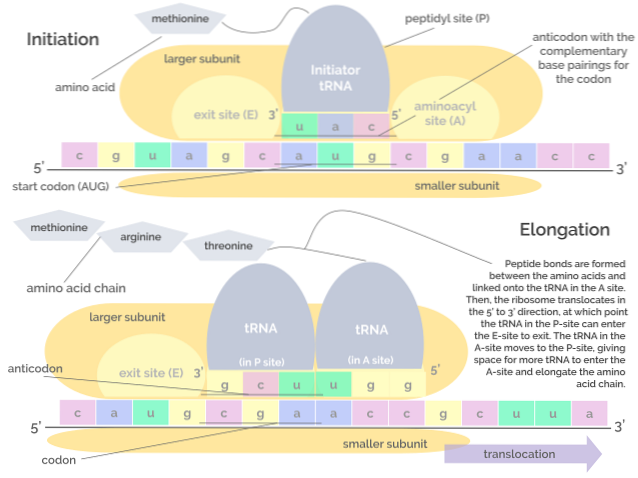
Denne fasen består av "bevegelsen" av ribosomet langs mRNA-molekylet og oversettelsen av hvert kodon som "leser", noe som innebærer vekst eller forlengelse av polypeptidkjeden ved fødselen..
Denne prosessen krever en faktor kjent som forlengelsesfaktor G og energi i form av GTP, som er det som driver translokasjonen av forlengelsesfaktorer langs mRNA-molekylet mens det blir oversatt..
Peptidyltransferaseaktiviteten til ribosomale RNAer tillater dannelse av peptidbindinger mellom suksessive aminosyrer som tilsettes kjeden.
Oversettelse slutter når ribosomet møter noen av termineringskodonene, siden tRNA ikke gjenkjenner disse kodonene (de koder ikke aminosyrer). Proteiner kjent som frigjøringsfaktorer binder også, noe som letter løsningen av mRNA fra ribosomet og dissosiasjonen av underenhetene..
I prokaryoter, som i eukaryote celler, finnes ribosomene som er ansvarlige for proteinsyntese i cytosolen (noe som også gjelder transkripsjonsmaskineriet), et faktum som tillater rask økning i den cytosoliske konsentrasjonen av et protein når ekspresjonen av gener som koder for det øker.
Selv om det ikke er en ekstremt vanlig prosess i disse organismer, kan de primære mRNAene som produseres under transkripsjon gjennomgå post-transkripsjonsmodning gjennom "spleising". Imidlertid er det vanligste å observere ribosomer festet til det primære transkriptet som oversetter det samtidig som det blir transkribert fra den tilsvarende DNA-sekvensen..
I lys av det ovennevnte begynner oversettelse i mange prokaryoter ved 5'-enden, siden 3'-enden av mRNA forblir festet til mal-DNA (og forekommer samtidig med transkripsjon)..
Prokaryote celler produserer også mRNA med utranslaterte regioner kjent som "Shine-Dalgarno-boksen" og hvis konsensus-sekvens er AGGAGG. Som det er tydelig er UTR-regionene til bakterier betydelig kortere enn de for eukaryote celler, selv om de utøver lignende funksjoner under translasjon..
I bakterier og andre prokaryote organismer er oversettelsesprosessen ganske lik den for eukaryote celler. Den består også av tre faser: initiering, forlengelse og avslutning, som avhenger av spesifikke prokaryote faktorer, forskjellige fra de som brukes av eukaryoter..
Forlengelse avhenger for eksempel av forlengelsesfaktorer kjent som EF-Tu og EF-Ts, snarere enn G-faktoren til eukaryoter..



Ingen har kommentert denne artikkelen ennå.