De Ribosomalt RNAeller ribosomal, i cellebiologi, er den viktigste strukturelle komponenten i ribosomer. Av denne grunn har de en uunnværlig rolle i syntesen av proteiner og er de mest vanlige i forhold til de andre hovedtypene av RNA: messenger og transfer..
Proteinsyntese er en avgjørende begivenhet i alle levende organismer. Tidligere ble det antatt at ribosomalt RNA ikke var aktivt involvert i dette fenomenet, og bare spilte en strukturell rolle. I dag er det bevis for at RNA har katalytiske funksjoner og er den sanne katalysatoren for proteinsyntese.
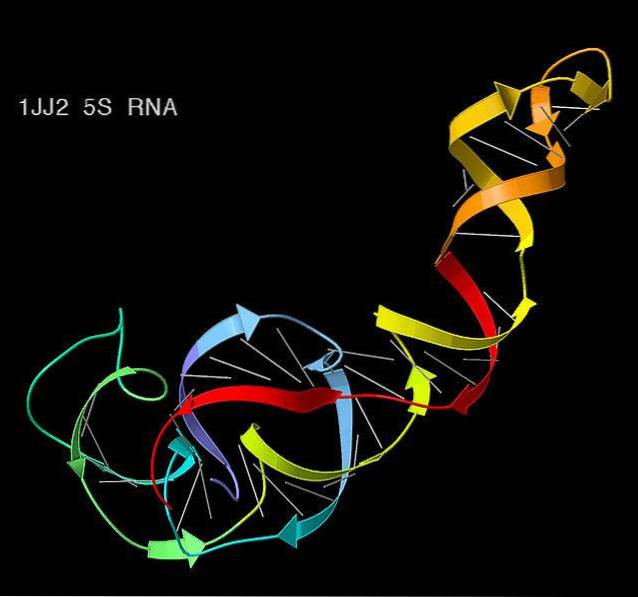
I eukaryoter er genene som gir opphav til denne typen RNA organisert i en region av kjernen som kalles nucleolus. RNA-typer klassifiseres vanligvis avhengig av deres oppførsel i sedimentering. Derfor blir de ledsaget av bokstaven S for "Svedberg-enheter".
Artikkelindeks
En av de mest slående forskjellene mellom eukaryote og prokaryote linjer er sammensetningen av ribosomalt RNA som utgjør deres ribosomer. Prokaryoter har mindre ribosomer, mens ribosomer i eukaryoter er større.
Ribosomer er delt inn i store og små underenheter. Det lille inneholder et enkelt ribosomalt RNA-molekyl, mens det store inneholder et større molekyl og to mindre, når det gjelder eukaryoter..
Det minste ribosomale RNA i bakterier kan være 1500 til 3000 nukleotider. Hos mennesker når ribosomalt RNA større lengder, mellom 1800 og 5000 nukleotider.
Ribosomer er de fysiske enhetene der proteinsyntese forekommer. De er sammensatt av omtrent 60% ribosomalt RNA. Resten er proteiner.
Historisk er ribosomalt RNA identifisert av sedimenteringskoeffisienten av suspenderte partikler sentrifugert under standardbetingelser, som er betegnet med bokstaven S for "Svedberg-enheter.".
En av de interessante egenskapene til denne enheten er at den ikke er additiv, det vil si at 10S pluss 10S ikke er 20S. Av denne grunn er det noe forvirring knyttet til den endelige størrelsen på ribosomene..
I bakterier, archaea, mitokondrier og kloroplaster inneholder den lille enheten i ribosomet 16S ribosomalt RNA. Mens den store underenheten inneholder to arter av ribosomalt RNA: 5S og 23S.
I eukaryoter finnes derimot 18S ribosomalt RNA i den lille underenheten, og den store underenheten, 60S, inneholder tre typer ribosomalt RNA: 5S, 5.8S og 28S. I denne avstamningen har ribosomer en tendens til å være større, mer komplekse og rikere enn i prokaryoter..
Ribosomalt RNA er den sentrale komponenten i ribosomer, så syntese er en uunnværlig hendelse i cellen. Syntese finner sted i kjernen, en region i kjernen som ikke er avgrenset av en biologisk membran.
Maskineriet er ansvarlig for å montere ribosomenhetene i nærvær av visse proteiner.
De ribosomale RNA-gener er organisert på forskjellige måter avhengig av avstamning. Husk at et gen er et segment av DNA som koder for en fenotype.
Når det gjelder bakterier, er genene for 16S, 23S og 5S ribosomale RNA organisert og transkribert sammen i et operon. Denne "gener sammen" -organisasjonen er veldig vanlig i prokaryote gener..
I kontrast er eukaryoter, mer komplekse organismer med en membranavgrenset kjerne, organisert i tandem. Hos oss mennesker er genene som koder for ribosomalt RNA organisert i fem “klynger” plassert på kromosomene 13, 14, 15, 21 og 22. Disse regionene kalles NOR.
I cellen er RNA-polymerase et enzym som er ansvarlig for å tilsette nukleotider til RNA-tråder. De danner et molekyl av disse fra et DNA-molekyl. Denne prosessen med dannelse av et RNA etter en DNA-herding er kjent som transkripsjon. Det finnes forskjellige typer RNA-polymeraser.
Generelt blir transkripsjonen av ribosomale RNA utført av RNA-polymerase I, med unntak av 5S ribosomalt RNA, hvis transkripsjon utføres av RNA-polymerase III. 5S har også den særegenheten at den transkriberes utenfor kjernen.
Promotorene for RNA-syntese består av to elementer som er rike på GC-sekvenser og en sentral region, her begynner transkripsjonen.
Hos mennesker binder transkripsjonsfaktorene som er nødvendige for prosessen til den sentrale regionen og gir opphav til pre-initieringskomplekset, som består av TATA-boksen og TBP-assosierte faktorer..
Når alle faktorene er sammen, binder RNA-polymerase I sammen med andre transkripsjonsfaktorer til den sentrale regionen av promoteren for å danne initieringskomplekset..
Senere skjer det andre trinnet i transkripsjonsprosessen: forlengelse. Her forekommer transkripsjonen og involverer tilstedeværelsen av andre katalytiske proteiner, som topoisomerase..
I eukaryoter har transkripsjonsenhetene til ribosomale gener en DNA-sekvens i 3'-enden med en sekvens kjent som Sal-boksen, som indikerer slutten på transkripsjonen.
Etter at transkripsjonen av de tandemordnede ribosomale RNAene har funnet sted, finner biogenesen av ribosomene sted i kjernen. Ribosomale genutskrifter modnes og assosieres med proteiner for å danne ribosomale enheter.
Før avslutning oppstår dannelsen av en serie "riboproteiner". Som i messenger RNA, er prosessen med skjøting drives av små nukleolære ribonukleoproteiner eller snRNP.
De skjøting det er en prosess der introner (ikke-kodende sekvenser) som vanligvis "avbryter" eksoner (sekvenser som koder for det aktuelle genet) blir eliminert.
Prosessen fører til mellomprodukter av 20S som inneholder 18S rRNA og 32S, som inneholder 5.8S og 28S rRNA.
Etter at ribosomale RNA har oppstått, gjennomgår de ytterligere modifikasjoner. Disse involverer metyleringer (tilsetning av en metylgruppe) på ca. 100 nukleotider per ribosom ved 2'-OH-gruppen i ribosomet. I tillegg forekommer isomerisering av mer enn 100 uridiner til pseudo-uridinformen..
I likhet med DNA er RNA sammensatt av en nitrogenholdig base kovalent bundet til en fosfat ryggrad..
De fire nitrogenholdige basene som danner dem er adenin, cytosin, uracil og guanin. Imidlertid, i motsetning til DNA, er RNA ikke et dobbeltbåndsmolekyl, men et enkelt bånd..
I likhet med overførings-RNA er ribosomalt RNA preget av å ha en ganske kompleks sekundær struktur, med spesifikke bindingsregioner som gjenkjenner messenger-RNA og overførings-RNA..
Hovedfunksjonen til ribosomalt RNA er å gi en fysisk struktur som gjør at messenger RNA kan tas og dekodes til aminosyrer for å danne proteiner..
Proteiner er biomolekyler med et bredt spekter av funksjoner - fra transport av oksygen, for eksempel hemoglobin, til støttefunksjoner.
Ribosomalt RNA brukes mye, både innen molekylærbiologi og evolusjon, så vel som i medisin..
Hvis du vil vite fylogenetiske forhold flere problemer mellom to grupper av organismer - det vil si hvordan organismer er relatert til hverandre, når det gjelder slektskap - blir ribosomale RNA-gener ofte brukt som tag.
De er veldig nyttige som molekylære markører takket være deres lave evolusjonshastigheter (disse typer sekvenser er kjent som "konserverte sekvenser").
Faktisk ble en av de mest kjente fylogenetiske rekonstruksjonene innen området biologi utført av Carl Woese og kolleger ved bruk av 16S ribosomale RNA-sekvenser. Resultatene av denne studien tillot levende organismer å bli delt inn i tre domener: archaea, bakterier og eukaryoter..
På den annen side er ribosomalt RNA ofte målet for mange antibiotika som brukes i medisin for å kurere et bredt spekter av sykdommer. Det er logisk å anta at ved å angripe proteinproduksjonssystemet til en bakterie, vil den umiddelbart bli påvirket..
Det er spekulert i at ribosomer, slik vi kjenner dem i dag, begynte dannelsen i svært avsidesliggende tider, nær dannelsen av LUCA (for dets akronym i Engelsk siste universelle felles forfedre eller siste universelle felles forfedre).
Faktisk sier en av hypotesene om livets opprinnelse at livet stammer fra et RNA-molekyl - siden det har de nødvendige autokatalytiske evnene for å bli betraktet som et av forløpsmolekylene i livet..
Forskerne foreslår at de nåværende ribosomforløperne ikke var like selektive med aminosyrer, og aksepterte både l- og d-isomerer. I dag er det allment kjent at proteiner dannes utelukkende fra aminoform l.
I tillegg har ribosomalt RNA evnen til å katalysere peptidyltransferasreaksjonen. Denne egenskapen til å tjene som et nukleotidregister, kombinert med sine katalytiske evner, gjør det til et nøkkelelement i utviklingen av de første formene på jorden.


Ingen har kommentert denne artikkelen ennå.