
De autofagi det er et intracellulært nedbrytningssystem som forekommer på en konservert måte i lysosomene til alle eukaryote celler (og gjærens vakuoler). Ordet brukes vanligvis for å referere til nedbrytning av komponentene i cytosolen eller "delene" av cellen som er "foreldet" eller som har sluttet å fungere ordentlig..
Begrepet autophagy ble laget i 1963 ved Rockefeller University av de Duve, som også observerte og beskrev prosessene med cellulær endocytose. Bokstavelig talt betyr ordet autophagy "å konsumere seg selv", selv om noen forfattere beskriver det som en "selvkanibalisme".
Dette systemet skiller seg fra proteasom-mediert nedbrytning ved at autofagi er i stand til å fjerne komplette intracellulære organeller og store proteinkomplekser eller aggregater på en ikke-selektiv måte..
Til tross for denne ikke-selektive fagocytosen har forskjellige undersøkelser vist at autofagi har mange fysiologiske og patologiske implikasjoner. Siden det aktiveres i perioder med tilpasning til sult, under utvikling, for eliminering av invaderende mikroorganismer, under programmert celledød, for eliminering av svulster, presentasjon av antigener, etc..
Artikkelindeks
Autophagy, som diskutert, er en prosess mediert av en cytoplasmatisk organell kjent som lysosomet..
Prosessen med "autophagy" begynner med innkapslingen av organellen som vil bli nedbrutt av en dobbel membran, og danner en membranaktig kropp kjent som autophagosome. Autofagosommembranen smelter deretter sammen med den lysosomale membranen eller med et sent endosom.
Hvert av disse trinnene mellom sekvestrering, nedbrytning og frigjøring av aminosyrer eller andre komponenter for resirkulering, utøver forskjellige funksjoner i forskjellige cellulære sammenhenger, noe som gjør autofagi til et svært multifunksjonelt system..
Autophagy er en ganske kontrollert prosess, siden bare de merkede mobilkomponentene er rettet mot denne nedbrytingsveien, og merkingen vanligvis skjer under cellulære ombyggingsprosesser..
For eksempel når en levercelle etablerer en avgiftningsrespons som respons på fettløselige medikamenter, prolifererer det glatte endoplasmatiske retikulumet betydelig, og når stimulansen generert av medikamentet avtar, fjernes det overflødige glatte endoplasmatiske retikulumet fra det cytosoliske rommet ved autofagi..
En av hendelsene som oftest utløser autofagiske prosesser er sult..
Avhengig av organismen som vurderes, kan forskjellige typer essensielle næringsstoffer utløse dette "resirkuleringssystemet". I gjær, for eksempel, selv om mangelen på karbon i visse aminosyrer og nukleinsyrer kan indusere autofagi, er mangelen på nitrogen den mest effektive stimulansen, som også gjelder for planteceller.
Selv om det ikke er helt forstått, har celler spesielle "sensorer" for å bestemme når et næringsstoff eller essensiell aminosyre er i veldig lav tilstand, og dermed utløse hele resirkuleringsprosessen gjennom lysosomer.
Hos pattedyr deltar noen hormoner i reguleringen (positiv eller negativ) av autofagi i celler som tilhører visse organer, som insulin, noen vekstfaktorer eller interleukiner, etc..
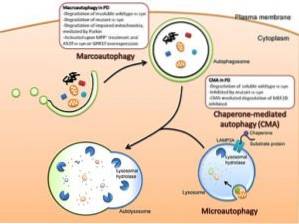
Det er tre hovedtyper av autophagy blant eukaryoter: macro autophagy, micro autophagy, og chaperone-mediert autophagy. Med mindre spesifisert, refererer begrepet autophagy til makro autophagy..
Selv om de tre typene autofagi er morfologisk forskjellige, ender de alle i transport av stoffer til lysosomer for nedbrytning og resirkulering..
Dette er en type autofagi som avhenger av treningen de novo av fagocytiske vesikler kjent som autofagosomer. Dannelsen av disse vesiklene er uavhengig av dannelsen av membran “knopper”, siden de dannes ved ekspansjon.
I gjær begynner dannelsen av autofagosomer på et bestemt sted kjent som PAS, mens det hos pattedyr forekommer mange forskjellige steder i cytosolen, sannsynligvis knyttet til det endoplasmatiske retikulumet gjennom strukturer kjent som "omegasomer".
Størrelsen på autofagosomer er svært variabel og avhenger av organismen og typen molekyl eller organell som er fagocytosert. Det kan variere fra 0,4-0,9 mikrometer i diameter i gjær til 0,5-1,5 mikrometer hos pattedyr..
Når membranene i autofagosomet og lysosomet smelter sammen, blandes innholdet av disse, og det er da fordøyelsen av målsubstratene til autofagi begynner. Denne organellen er da kjent som autolysosomet..
For noen forfattere kan makroautofagi subklassifiseres i sin tur til indusert autofagi og baseline autofagi. Indusert makroautofagi brukes til å produsere aminosyrer etter en lengre periode med sult.
Basal makroautofagi refererer til den konstitutive mekanismen (som alltid er aktiv) som er viktig for omsetningen av de forskjellige cytosoliske komponentene og intracellulære organeller.
Denne typen autofagi refererer til prosessen der det cytoplasmatiske innholdet blir introdusert til lysosomet gjennom invaginasjoner som forekommer i organellens membran..
Når de er introdusert i lysosomet, flyter vesiklene produsert av disse invaginasjonene fritt i lumenet til de lyseres og innholdet frigjøres og nedbrytes av spesifikke enzymer..
Denne typen autofagi er bare rapportert for pattedyrceller. I motsetning til makro-autofagi og mikro-autofagi, hvor noen cytosoliske deler er ikke-spesifikt fagocytosert, er autofagi formidlet av kaperoner ganske spesifikk, siden det avhenger av tilstedeværelsen av spesielle pentapeptidsekvenser i substratene som vil bli fagosytosert..
Noen forskere har bestemt at dette pentapeptidmotivet er relatert til KFERQ-sekvensen, og at det finnes i mer enn 30% av cytosolproteiner..
Det kalles "chaperonemediert" siden chaperoneproteiner er ansvarlige for å holde dette konserverte motivet eksponert for å lette dets gjenkjenning og forhindre at proteinet brettes på det..
Proteiner med denne merkelappen blir translokert til lysosomal lumen og der blir de nedbrutt. Mange av nedbrytningssubstratene er glykolytiske enzymer, transkripsjonsfaktorer og deres hemmere, kalsium- eller lipidbindende proteiner, proteasom-underenheter og noen proteiner involvert i vesikulær handel..
Som de to andre typene autofagi, er chaperonemediert autofagi en regulert prosess på mange nivåer, fra gjenkjenning av etiketter til transport og nedbrytning av substrater i lysosomer..
En av hovedfunksjonene til den autofagiske prosessen er eliminering av senescent eller "foreldede" organeller, som er merket av forskjellige veier for deres nedbrytning i lysosomer..
Takket være observasjonen av elektronmikrofotografier av lysosomer i pattedyrceller, har tilstedeværelsen av peroksisomer og mitokondrier blitt oppdaget i dem..
I en levercelle er for eksempel den gjennomsnittlige levetiden til en mitokondrion 10 dager, hvoretter denne organellen blir fagocytosert av lysosomer, der den nedbrytes og komponentene resirkuleres for forskjellige metabolske formål..
Under forhold med lav næringskonsentrasjon kan celler utløse dannelsen av autofagosomer for selektivt å "fange" deler av cytosolen, så vel som de fordøyde metabolittene i disse autofagosomene kan hjelpe celler til å overleve når ytre forhold er begrensende fra synspunkt. ernæringsmessig synspunkt.
Autophagy har viktige funksjoner i restrukturering av celler i differensieringsprosessen, siden det deltar i forkastning av cytosoliske deler som ikke er nødvendige på bestemte tidspunkter..
Det har også viktige implikasjoner for cellehelsen, da det er en del av forsvarsmekanismene mot invaderende virus og bakterier..
Yoshinori Ohsumi, en Nobelprisvinnende japansk forsker i fysiologi og medisin i 2016, beskrev de molekylære mekanismene for autofagi i gjær mens han studerte den metabolske skjebnen til mange proteiner og vakuolene til disse encellede organismer..
I sitt arbeid identifiserte Ohsumi ikke bare proteinene og banene som var involvert i prosessen, men demonstrerte også hvordan autofagveien reguleres takket være virkningen av proteiner som er i stand til å "telle" forskjellige metabolske tilstander..
Arbeidet deres begynte med nøyaktige mikroskopiske observasjoner av vakuoler under intense nedbrytningshendelser. Vakuoler regnes som lagringsplasser for gjær "søppel" og mobilavfall..
Ved å observere gjær med defekte mutante genotyper for forskjellige gener relatert eller hypotetisk relatert til autofagi (kjent som genene ATG), var denne forskeren og hans samarbeidspartnere i stand til å beskrive det autofagiske gjærsystemet på genetisk nivå.
Deretter bestemte denne gruppen forskere de viktigste genetiske egenskapene til proteinene kodet av disse genene og ga betydelige bidrag om deres interaksjon og dannelsen av kompleksene som var ansvarlige for initiering og utførelse av autofagi i gjær..
Takket være arbeidet til Yoshinori Ohsumi, forstår vi i dag de molekylære aspektene av autophagy, så vel som viktige implikasjoner for riktig funksjon av cellene og organene som komponerer oss..



Ingen har kommentert denne artikkelen ennå.