
CD3, I immunologi er de akronymer som beskriver "gruppen av differensiering 3" (Cluster of Differentiering 3) og definerer et proteinkompleks som er tilstede i plasmamembranen til celler i immunsystemet kjent som T-lymfocytter..
Proteinene i CD3-komplekset assosieres normalt med et annet proteinkompleks i plasmamembranen til lymfocytter kalt T-celle reseptor eller TCR (T Cell Receptor)..
TCR er en heterodimer sammensatt av to peptidkjeder bundet sammen av disulfidbindinger. Som det kan utledes av navnet, er TCR unik for celler av T-lymfocyttlinjen og har viktige implikasjoner for immunforsvaret til disse cellene..
I tillegg har hver T-celle en spesifikk TCR, siden disse proteinene faktisk er et slags antistoff, så de er i stand til å gjenkjenne bare en type definert antigen..
Proteinene i CD3-komplekset har transcendentale funksjoner i transduksjon av signaler relatert til interaksjonen mellom TCR-komplekset og dets spesifikke antigen, som de deltar i en viktig del av utviklingen av T-lymfocytter kjent som "aktivering".
Artikkelindeks
CD3 blir av mange forfattere betraktet som et "coreceptor" -kompleks av T-celle-reseptorkomplekset (TCR). Det er et molekyl som uttrykkes i de tidlige stadiene av T-lymfocyttutvikling..
Den er tilstede i både hjelper-T-lymfocytter ("hjelpere") og cytotoksiske T-lymfocytter, men har ikke blitt påvist i andre lymfoide celler som B-celler eller naturlige killer (NK) -celler..
CD3-komplekset er et proteinkompleks av fem uforanderlige polypeptidkjeder kjent som γ, ε, δ, ζ og η; Disse kjedene assosieres med hverandre for å danne tre dimere strukturer: γε heterodimeren, δε heterodimeren og ζζ homodimeren eller heterη heterodimeren.
90% av CD3-kompleksene har hom-homodimeren, mens ζη-heterodimeren bare er funnet i de resterende 10%..
Med unntak av chains og η-kjedene, er peptidkjedene til CD3-komplekset kodet av forskjellige gener. ζ og η er kodet av det samme genet, men går gjennom forskjellige skjøtingsprosesser (“spleising”).
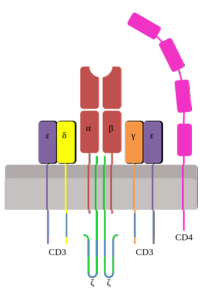
Γ-, ε- og δ-proteinene til CD3-komplekset er en del av immunoglobulin-superfamilien og er transmembrane proteiner. De har et transmembrandomene, et cytosolisk domene på mer enn 40 aminosyrer og et ekstracellulært domene (immunoglobulin-type).
Peptidkjeden ζ er ganske forskjellig fra de andre tre: dens ekstracellulære del er ca. 9 aminosyrer lang, har et kort transmembransegment og har et cytosolisk domene på 113 aminosyrer..
Peptidkjedene til CD3-komplekset har en transmembranregion som har en asparaginsyre eller glutaminsyrerest (negativt ladede rester), som er i stand til å samhandle med de positive ladningene av aminosyrene i den transmembrane delen av TCR-komplekset..
Co-mottaksfunksjonen til CD3-komplekset med TCR-komplekset er nært knyttet til den "transmembrane" interaksjonen av restene av polypeptidkjedene som utgjør begge kompleksene..
Alle cytosoliske kjeder av CD3-komplekset har et tyrosinbasert immunreseptoraktiveringsmotiv (ITAM)..
Disse ITAM-motivene er ansvarlige for signaltransduksjon innover, da de samhandler med tyrosinkinaseenzymer, som er viktige formidlere i intracellulær signalering..
Å vite at CD3 er et kompleks av flere komponenter, er det viktig å spesifisere at det samarbeider med interaksjonen av antigenet med TCR-reseptoren, men ikke deltar i det, det vil si at det ikke kommer i kontakt med antigenet..
Mange bevislinjer antyder at CD3 ikke bare er nødvendig for å formidle antigen-antistoffinteraksjon på overflaten av T-celler, men at dets ekspresjon er nødvendig for ekspresjonen av selve TCR-komplekset..
Den ekstracellulære delen av CD3-komplekset brukes som et "antigen" for antistoffgjenkjenning av celler av T-lymfocyttlinjen, noe som er viktig med tanke på klinisk cytologi og sykdomsdiagnose..
T-celler eller lymfocytter deltar i de viktigste fenomenene i den humorale og cellulære immunresponsen, som avhenger betydelig av deres aktivering og multiplikasjon.
CD3-molekylkomplekset virker under T-celleaktivering ved å samhandle med TCR-komplekset og danne TCR-CD3 "effektor" -komplekset..
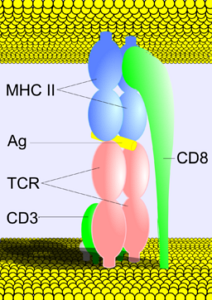
Husk at dannelsen av dette komplekset bare vil skje når den aktuelle T-cellen gjenkjenner et antigen som presenteres for det i sammenheng med et molekyl av hovedhistokompatibilitetskomplekset eller MHC (Major Histocompatibility Complex) i klasse I eller klasse II, avhengig av på typen T-lymfocytt.
Antigen-MHC / TCR kompleks / CD3 kompleks interaksjon utløser en komplisert signalprosess, som begynner i T-lymfocyttmembranen og ender i cellekjernen med stimulering av transkripsjonen av spesifikke gener involvert i cellesyklusen og differensiering..
CD3, som sagt, samarbeider om signaltransduksjon, siden ITAM-domenene til dets polypeptidkjeder samhandler med en kaskade av proteintyrosinkinaser som for det meste aktiveres ved fosforylering..
Tyrosinkinaseenzymer "rekrutterer" og aktiverer andre elementer nedstrøms i signalkjeden, spesielt noen "stillas" proteiner og andre enzymer med evnen til å aktivere eller indusere frigjøring av molekyler som fungerer som andre budbringere og transkripsjonsfaktorer..
I tillegg til hva som skjer på stedene for interaksjon og utveksling av signaler mellom to nevroner (neuronale synapser), har interaksjonssidene mellom celler som presenterer antigener i sammenheng med MHC-molekyler og membranreseptorer på overflaten av nevroner. T-lymfocytter har vært betegnet "immnosinapsis".
CD3-komplekset, siden det deltar aktivt i interaksjonen, er en viktig del av immunsynapsisidene.


Ingen har kommentert denne artikkelen ennå.