De NK-celler (av engelsk Natural Killerceller), naturlige drapsceller eller naturlige cytocidale celler, er en type effektorlymfocytt som deltar i responsene til det medfødte eller uspesifikke immunsystemet.
Disse cellene ble oppdaget for mer enn 40 år siden, og noen forfattere beskriver dem som "granulære lymfocytter" som, i motsetning til T- og B-lymfocytter, deltar i den medfødte immunresponsen og ikke gjennomgår genetiske omleggingsprosesser i deres kimlinjer..
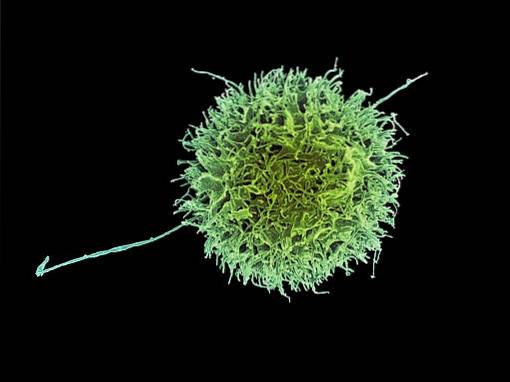
Siden de ikke uttrykker de vanlige markørene for de to andre klassene lymfocytter, ble NK-celler opprinnelig kalt "nullceller". Imidlertid viste videre studier at de var lymfocytter med store granulocytter..
Disse cellene er i stand til å kontrollere forskjellige typer svulster og mikrobielle infeksjoner ved å begrense spredning og vevskader. Videre kan de lyse forskjellige typer celler uten definert antigen stimulering..
NK-celler er ekstremt viktige celler i første forsvarslinje mot patogener, et faktum som har blitt vist gjennom studier der mennesker med mangel på NK-celler kan lide dødelige infeksjoner i barndommen..
Artikkelindeks

NK-celler finnes i en lavere andel enn noen av de to andre klassene lymfocytter (de utgjør 2 til 10% av sirkulerende lymfocytter), og siden de tilhører det medfødte forsvarssystemet, antas det at de var blant de første cellulære elementene involvert i beskyttelse av flercellede organismer.
I likhet med T-lymfocytter og B-lymfocytter er NK-celler en del av pattedyrets hematopoietiske system og er avledet fra stamfarhematopoietiske celler som uttrykker CD34 + membranmarkører, som også er kjent som HPC-celler..
Mens T-celler er kjent for å modnes i thymus og B-celler for å modnes i benmargen, har forsøk på å bestemme den komplette utviklingsveien til NK fra HPC-forløpere ikke vært vellykket; bare kjent for å være thymusuavhengig.
NK-celler uttrykker adhesjonsmolekyler på membranoverflaten kjent som CD2, LFA-1, NCAM eller CD56. De uttrykker også reseptorer med lav affinitet til den konstante delen (Fc) av immunglobulin IgG som kollektivt kalles FcyRIIIA eller CD16..
Det indre av en naturlig cytocidcelle er fullpakket med store cytosoliske granuler som er fylt med perforin, granzymer og proteoglykaner..
Perforiner er poredannende proteiner som "gjennomborer" plasmamembranen til celler som blir angrepet av NK-er. Granzymes, derimot, er serinproteaser som tar seg inn i celler gjennom porene dannet av perforiner og nedbryter intracellulære proteiner..
Den kombinerte virkningen av perforiner og granzymer resulterer i arrestasjonen av produksjonen av virale eller bakterielle proteiner og i apoptose eller programmert celledød av den infiserte cellen.
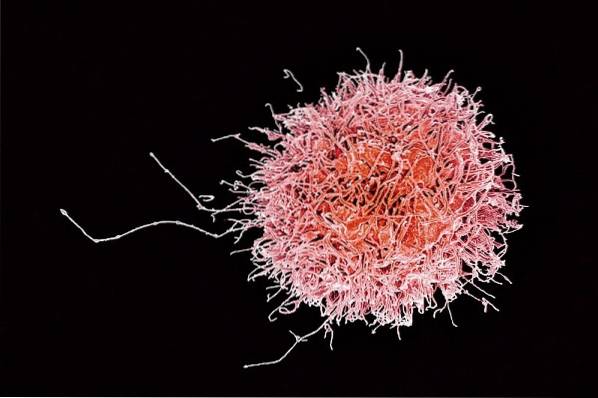
Naturlige drapsceller fungerer i eliminering av "mål" eller "mål" celler på en naturlig måte, det vil si spontant og uten mye spesifisitet, siden de ikke krever noen form for antigen priming..
En av de viktigste funksjonene til denne gruppen celler er dens evne til å drepe tumorceller, spesielt de som tilhører hematopoietiske linjer, samt celler som er invadert av forskjellige typer virus og / eller bakterier..
Aktiviteten stimuleres sterkt av faktorer som IFN-α og β interferoner, så vel som av interleukin IL-12..
Takket være at disse cellene produserer noen viktige cytokiner for immunforsvaret, deltar NK i immunregulering, både i det medfødte og adaptive eller spesifikke systemet..
For eksempel kan produksjonen av interferon gamma (IFN-γ) i NK-celler forstyrre deltakelsen av makrofager i medfødt immunitet, da dette molekylet forstyrrer fagocytiske og mikrobicide aktiviteter.
Samtidig kan IFN-γ produsert av naturlige cytocider endre forpliktelsen til hele populasjoner av hjelper-T-celler, siden IFN-γ også hemmer utvidelse og utvikling av en populasjon i forhold til en annen..
NK-celler representerer den første forsvarslinjen under virusinfeksjoner, ettersom de kontrollerer replikering av virus mens cytotoksiske T-celler aktiveres, sprer seg og differensierer, noe som kan ta mer enn 6 dager.
NK-cellepopulasjoner er ganske heterogene, både fenotypisk, funksjonelt og anatomisk. I tillegg avhenger dens egenskaper av hvilken type organisme som studeres..
I den murine (musemodellen) er det beskrevet tre forskjellige sett med naturlige cytocidale celler som skiller seg fra hverandre ved uttrykk for markørene CD11b og CD27. I denne forstand er det celler CD11bdullCD27 +, CD11b + CD27 + og CD11b + CD27dull.
Overskriftet "kjedelig" refererer til "av" eller "inaktivt" og brukes, i dette tilfellet, til å beskrive dets tilstand på overflaten av murine celler..
CD11bdull CD27 + celler skiller seg fra en dobbel positiv type forløper (CD11b + CD27 +) som igjen gir opphav til den mer modne typen NK-celler hos gnagere: CD11b + CD27dull.
Både de doble positive linjene og CD11b + CD27 kjedelige linjene er preget av å eliminere målcellene og ved å utskille et cytokin kjent som interferon (INF-γ). Imidlertid er sistnevnte i noe som kalles "replikativ senescens".
De tre typene NK-celler er fordelt i forskjellige vev. CD11bdull CD27 + celler er hovedsakelig i lymfeknuter og benmarg. CD11b + CD27 kjedelige celler er rikelig i blod, milt, lunger og lever; i mellomtiden har dobbeltpositive celler en mer homogen eller systemisk fordeling.
NK-celler hos mennesker er også klassifisert i henhold til overflatemarkørene de uttrykker, men i dette tilfellet blir de differensiert av tilstedeværelsen av markørene CD56dim og CD56bright. Håndskriftene "dim" og "lyst" refererer til henholdsvis "mørk" og "lys"..
Forskjellene mellom disse cellene ligger i "målsøk" -egenskapene til hver enkelt, som er gitt ved tilstedeværelsen av den ene eller den andre markøren..
I perifert blod og milt hos mennesker er hovedtypen av NK-celler kjent som CD56dimCD16 +, som vanligvis uttrykker porfyrinproteinet og er cytotoksisk. De produserer også IFN-γ som et resultat av interaksjon med tumorceller under forhold in vitro.
CD56 lys CD16-celler finnes i lymfeknuter og mandler, som i stedet for å produsere porfyrin, skiller ut cytokinet IFN-y som respons på stimulering av interleukiner IL-12, IL-15 og IL-18.
Hos mennesker og gnagere antas mandlene og andre sekundære lymfoide organer å være produksjonssteder og modning for de fleste NK-celler..
Noen studier antyder at det er en viss likhet mellom humane CD56bright-celler og CD11-kjedelige celler når det gjelder anatomisk plassering, fenotypiske egenskaper, cytosolisk perforininnhold, proliferativt potensial og overflateekspresjon av interleukin IL-7R..
Disse har ganske kort halveringstid (ca. 2 uker), og det antas at det i et voksent menneske er rundt 2 billioner celler i omløp. De er rikelig i blod, milt og annet lymfoide og ikke-lymfoide vev.
Studier viser at den normale konsentrasjonen hos voksne menn og kvinner er rundt 200 og 600 celler per mikroliter testet blod..
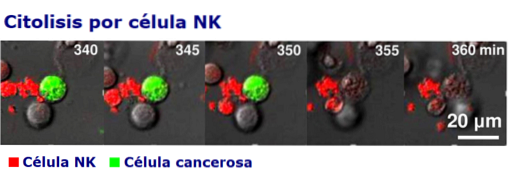
Intensiteten og kvaliteten på de cytotoksiske responsene til NK-celler avhenger av mikromiljøet som genereres av cytokinene og av interaksjonen med andre celler i immunsystemet, spesielt med T-celler, dendritiske celler og makrofager..
Blant de aktiverende cytokinene til NK-celler er interleukiner, spesielt IL-12, IL-18 og IL-15; samt type I interferon (IFN-I). Interferon og interleukiner er kraftige aktivatorer av NKs effektorfunksjon.
Interleukin IL-2 er også involvert i å fremme spredning, cytotoksisitet og cytokinsekresjon av NK-celler..
IL-15 er avgjørende for differensiering av NKs, mens IL-2 og IL-18 er essensielle for den påfølgende modningen av slike celler..
Naturlige cytocidale celler aktiveres takket være gjenkjenningen av egne molekyler (en prosess kjent på engelsk som “anerkjennelse av selvmolekyler”) Dette uttrykkes konstitusivt under stabile forhold.
I cellene deres uttrykker disse cellene forskjellige medlemmer av en familie av overflateproteiner som inneholder to eller tre immunglobulinlignende domener i deres ekstracellulære deler og motiver som ligner aktiveringsdomenene til immunreseptorer via tyrosin i deres intracellulære region..
Hver NK-celle kan uttrykke en eller flere av disse reseptorproteinene, og hver reseptor er i stand til å gjenkjenne en spesifikk form av et hovedhistokompatibilitetskompleks klasse I (MHC-I) molekyl..
Gjenkjennelsen mellom dette molekylet og reseptoren på overflaten av naturlige cytocidale celler fører til dannelsen av et kompleks med rikelig peptider avledet fra "selv" proteiner..
Reseptorene er for det meste inhiberende proteiner som aktiverer en tyrosinfosfatase som forhindrer at cellen avgir normale responser.
Eliminasjonen eller døden som medieres av naturlige drapsceller er lik den som oppstår under den cytolytiske virkningen av CD8 T-lymfocytter (cytotoksisk), selv om forskjellen er at NK er konstitutive cytotoksiske, det vil si at de ikke trenger å aktiveres før.
Aktive NK-er uttrykker FasL-liganden, og det er derfor de relativt lett induserer død av målceller som uttrykker Fas-proteinet på overflaten..
Etter dannelsen av den komplette FasL / Fas, oppstår en prosess kjent som "degranulering", som ender med frigjøring av porfyrin og granzymer på de intercellulære kontaktstedene..
Til tross for de nevnte likhetene, skiller NK seg fra cytotoksiske T-celleformidlede mekanismer ved at gjenkjenningen av deres målceller ikke er avhengig av proteiner fra det viktigste histokompatibilitetskomplekset..
En annen forskjell er at NK-celler ikke har et "immunminne" -system, noe som demonstreres av det faktum at deres aktivitet ikke øker etter en annen eksponering for målcellene..
Naturlige cytocider skiller mellom en sunn celle og en annen infisert eller svulstig (kreft) takket være en balanse mellom aktiverende og inhiberende signaler, som gjenkjennes av spesifikke overflatereseptorer.
Disse reseptorene er av to typer: lektintype (proteiner som binder karbohydrater og andre proteiner) og immunglobulintype (ligner på den konstante regionen av immunglobuliner).
I denne siste gruppen gjenkjennes drapscellene immunoglobulinreseptorer eller KIR. drapscelle-immunglobulinlignende reseptorer), som er i stand til gjenkjenning og binding av spesifikke former av proteinene i klasse I major histokompatibilitetskompleks (HLA-B eller HLA-C).
Det er viktig å merke seg at NK ikke "angriper" celler som uttrykker normale nivåer av MHC klasse I-molekyler, men de dreper celler som uttrykker fremmede molekyler av denne typen eller de som mangler nevnte markører (noe som er typisk i tumorceller og infiserte av virus).
NK-er uttrykker noen vanlige membranmarkører for monocytter og granulocytter, og andre som er typiske for T-lymfocytter..
På den annen side uttrykker naturlige cytocider forskjellige grupper av overflatemarkører, men det er foreløpig ikke kjent med sikkerhet om heterogeniteten indikerer celleundpopulasjoner eller stadier under aktivering eller modning..
Noen eksempler på NK-cellemarkører er:
NK-celler er avledet fra den samme forelderen som gir opphav til T-celler. Denne overordnede cellen uttrykker vanligvis markørene CD7, CD2 og noen ganger CD5..
CD2 er et 50 kDa molekylvektprotein som også er tilstede i T-celler. Det er kjent som et overflateadhesjonsmolekyl og deltar i aktivering av T-celler..
CD5 er normalt tilstede på T-celler og noen delpopulasjoner av B. Den er en 67 kDa markør og har også limfunksjoner..
CD7-markøren er typisk for hematopoietiske stamceller og har også blitt funnet i visse T-celle-subpopulasjoner. Den har en molekylvekt på 40 kDa og fungerer i signaltransduksjon..
Denne reseptoren deles mellom NK, monocytter og granulocytter. Den har en molekylvekt på 165 kDa og kan assosieres med andre overflatemarkører. Hovedfunksjonene er lim, spesielt under fagocytose eller "opsonisering" -prosesser..
Det er en 50-70 kDa reseptor som er bundet til et transmembranfosfatidylinositolmolekyl. Deltar i aktivering av naturlige drapsceller og finnes også i granulocytter og makrofager.
Den fungerer også som en reseptor for den konstante regionen av gammakjeden til noen antistoffer..
Den finnes på de fleste T-lymfocytter og er en 55 kDa peptidkjedehomodimer. Det ser ut til å være et medlem av tumornekrosefaktor (TNF-R) -reseptorfamilien og er også involvert i ko-stimulering av T-celler..
Denne reseptoren er unik for NK-celler og består av 135 og 220 kDa-kjeder. Deltar i den "homotypiske" vedheftingen av disse cellene.



Ingen har kommentert denne artikkelen ennå.