De erytropoietin, hemopoietin eller EPO er et glykoprotein med hormonelle funksjoner (cytokin) som er ansvarlig for kontrollen av spredning, differensiering og overlevelse av stamceller av erytrocytter eller røde blodlegemer i benmargen, det vil si av erytropoies.
Dette proteinet er en av de forskjellige vekstfaktorene som styrer de hematopoietiske prosessene, fra hvilke celler fra en liten gruppe pluripotente stamceller dannes: både erytrocytter og hvite blodlegemer og lymfocytter. Det vil si celler av myeloide og lymfoide linjer.
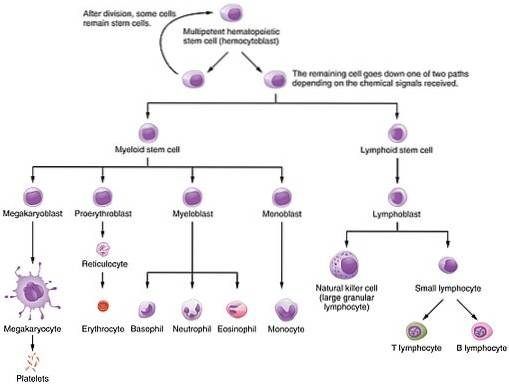
Dens betydning ligger i den funksjonelle betydningen av celler som hjelper til med å formere seg, differensiere og modne, siden erytrocytter er ansvarlige for transporten av oksygen fra lungene til kroppens forskjellige vev..
Erytropoietin var den første vekstfaktoren som ble klonet (i 1985), og administrasjonen av den for vellykket behandling av anemi forårsaket av nyresvikt er for tiden godkjent av American Food and Drug Administration (FDA)..
Forestillingen om at erytropoiesis kontrolleres av en humoral faktor (løselig faktor som er tilstede i sirkulasjonen) ble foreslått for mer enn 100 år siden av Carnot og Deflandre når man studerte de positive effektene på økningen av prosentandelen av røde blodlegemer hos kaniner behandlet med serum. dyr.
Det var imidlertid først i 1948 at Bonsdorff og Jalavisto introduserte begrepet "erytropoietin" for å beskrive den humorale faktoren med en spesifikk implikasjon på produksjonen av erytrocytter..
Artikkelindeks
Erytropoietin er et protein fra glykoproteinfamilien. Den er stabil ved sure pH-er og har en molekylvekt på omtrent 34 kDa..
Den har ca. 193 aminosyrer, inkludert en hydrofob N-terminal region med 27 rester, som fjernes ved co-translasjonell prosessering; og en argininrest i posisjon 166 som også går tapt, så det sirkulerende proteinet har 165 aminosyrer.
I sin struktur kan det sees dannelse av to disulfidbroer mellom cysteinrestene som er tilstede i posisjonene 7-161 og 29-33, som er knyttet til dens drift. Den består mer eller mindre enn 50% av alfahelices, som tilsynelatende deltar i dannelsen av en kuleformet region eller del.
Den har 40% karbohydrater, representert ved tre oligosakkaridkjeder N-koblet til forskjellige asparaginsyrerester (Asp), og en O-kjede knyttet til en serinrest (Ser). Disse oligosakkaridene består hovedsakelig av fukose, mannose, N-acetylglukosamin, galaktose og N-acetylneuramininsyre..
Karbohydratregionen i EPO oppfyller flere roller:
- Det er viktig for dets biologiske aktivitet.
- Beskytter den mot nedbrytning eller skade forårsaket av oksygenfrie radikaler.
- Oligosakkaridkjedene er nødvendige for utskillelsen av det modne proteinet.
Hos mennesker ligger genet som koder for dette proteinet midt i den lange armen av kromosom 7, i regionen q11-q22; den finnes i en enkelt kopi i en 5.4kb-region og har fem eksoner og fire introner. Homologistudier indikerer at sekvensen deler 92% identitet med andre primates og 80% med noen gnagere..
Under fosterutvikling produseres erytropoietin hovedsakelig i leveren, men det er fastslått at genet som koder for dette hormonet, i samme fase, også blir rikelig uttrykt i den midtre regionen i nyrene nefroner..
Etter fødselen, i det som kan betraktes som alle postnatale stadier, produseres hormonet hovedsakelig i nyrene. Spesielt av cellene i cortex og overflaten av nyrekroppene.
Leveren deltar også i produksjonen av erytropoietin i postnatale stadier, hvorfra omtrent 20% av det totale sirkulerende EPO-innholdet skilles ut..
Andre "ekstra renale" organer der erytropoietinproduksjon er blitt påvist inkluderer perifere endotelceller, vaskulære glatte muskelceller og insulinproduserende celler..
Noen EPO-sekresjonssentre er også kjent for å eksistere i sentralnervesystemet, inkludert hippocampus, cortex, hjerneendotelceller og astrocytter..
Produksjonen av erytropoietin styres ikke direkte av antall røde blodlegemer i blodet, men av tilførsel av oksygen i vevet. Mangel på oksygen i vevet stimulerer produksjonen av EPO og dets reseptorer i leveren og nyrene.
Denne hypoksimedierte aktivering av genuttrykk er produktet av aktiveringen av banen til en familie av transkripsjonsfaktorer kjent som hypoksiinduserbar faktor 1 (HIF-1). hypoksiinducerbar faktor 1).
Hypoksia induserer da dannelsen av mange proteinkomplekser som oppfyller forskjellige funksjoner i aktivering av erytropoietinekspresjon, og som binder seg direkte eller indirekte til faktorer som oversetter aktiveringssignalet til promotoren til EPO-genet, og stimulerer dets transkripsjon..
Andre stressfaktorer som hypoglykemi (lavt blodsukker), økning i intracellulært kalsium eller tilstedeværelsen av reaktive oksygenarter, utløser også HIF-1-banen..
Virkningsmekanismen til erytropoietin er ganske kompleks og avhenger hovedsakelig av dens evne til å stimulere forskjellige signalkaskader involvert i celleproliferasjon, som i sin tur er relatert til aktivering av andre faktorer og hormoner.
I en sunn voksen menneskekropp er det en balanse mellom produksjon og ødeleggelse av røde blodlegemer eller erytrocytter, og EPO deltar i å opprettholde denne balansen ved å erstatte erytrocyttene som forsvinner..
Når oksygenmengden som er tilgjengelig i vevene er veldig lav, øker ekspresjonen av genet som koder for erytropoietin i nyrene og leveren. Stimuleringen kan også gis av høye høyder, hemolyse, tilstander med alvorlig anemi, blødning eller langvarig eksponering for karbonmonoksid.
Disse forholdene genererer en tilstand av hypoksi, som får utskillelsen av EPO til å øke, et større antall røde blodlegemer blir produsert og brøkdelen av retikulocytter i omløp, som er en av stamcellene til erytrocytter, øker også..
I erytropoiesis er EPO primært involvert i spredning og differensiering av stamceller som er involvert i røde blodlegemer (erytrocytiske stamfarer), men det aktiverer også mitose i proerythroblaster og basofile erytroblaster, og akselererer også frigjøring av benmargsretikulocytter.
Det første nivået som proteinet fungerer på er forebygging av programmert celledød (apoptose) av forløpercellene dannet i benmargen, som det oppnår ved inhiberende interaksjon med faktorene som er involvert i denne prosessen..
Celler som reagerer på erytropoietin har en spesifikk reseptor for erytropoietin kjent som erytropoietinreseptoren eller EpoR. Når proteinet danner et kompleks med reseptoren, overføres signalet inne i cellen: mot kjernen.
Det første trinnet for signaloverføring er en konformasjonsendring som oppstår etter at proteinet binder seg til reseptoren, som samtidig er bundet til andre reseptormolekyler som aktiveres. Blant dem er Janus-tyrosinkinase 2 (Jack-2).
Blant noen av banene som aktiveres nedstrøms, etter at Jack-2 har formidlet fosforylering av tyrosinrester av EpoR-reseptoren, er MAP-kinase- og proteinkinase C-banen, som aktiverer transkripsjonsfaktorer som øker ekspresjonen av spesifikke gener.
Som mange hormonelle faktorer i organismer er erytropoietin ikke begrenset til en enkelt funksjon. Dette er belyst gjennom en rekke undersøkelser.
I tillegg til å fungere som en proliferasjons- og differensieringsfaktor for erytrocytter, som er essensielle for transport av gasser gjennom blodbanen, ser det ut til at erytropoietin oppfyller noen tilleggsfunksjoner, ikke nødvendigvis knyttet til aktivering av celleproliferasjon og differensiering.
Studier har antydet at EPO forhindrer celleskade, og selv om dens virkningsmekanismer ikke er kjent, antas det at det kan forhindre apoptotiske prosesser produsert av redusert eller fraværende oksygenspenning, opphisse toksisitet og eksponering for frie radikaler..
Deltakelsen i forebygging av apoptose har blitt studert ved interaksjon med avgjørende faktorer i signalkaskadene: Janus-tyrosinkinase 2 (Jak2), caspase 9, caspase 1 og caspase 3, glykogensyntasekinase-3β, aktiveringsfaktor for apoptotiske proteaser 1 (Apaf-1) og andre.
Deltar i inhiberingen av cellebetennelse ved å hemme noen proinflammatoriske cytokiner som interleukin 6 (IL-6), tumornekrosefaktor alfa (TNF-α) og monocytt-kjemotrekkende protein 1.
I det vaskulære systemet har det vist seg å samarbeide om å opprettholde dets integritet og i dannelsen av nye kapillærer fra eksisterende kar i områder uten vaskulatur (angiogenese). I tillegg forhindrer det permeabiliteten til blod-hjerne-barrieren under skader.
Det antas å stimulere postnatal neovaskularisering ved å øke mobilisering av stamceller fra benmargen til resten av kroppen..
Det spiller en viktig rolle i utviklingen av nevrale stamceller gjennom aktivering av kjernefaktoren KB, som fremmer produksjonen av nervestamceller..
EPO fungerer sammen med andre cytokiner, og har en "modulerende" funksjon i kontrollen av spredning og differensieringsveier for megakaryocytter og granulocytt-monocytter..


Ingen har kommentert denne artikkelen ennå.