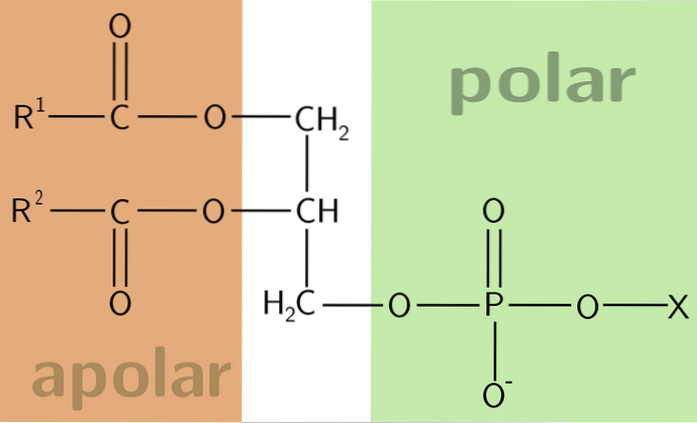
De fosfoglyserider eller glyserofosfolipider er lipidmolekyler som er rikelig i biologiske membraner. Et fosfolipidmolekyl består av fire basiske komponenter: fettsyrer, et skjelett festet til fettsyren, et fosfat og en alkohol festet til sistnevnte..
Generelt er det på karbon 1 av glyserol en mettet fettsyre (bare enkeltbindinger), mens på karbon 2 er fettsyren av den umettede typen (dobbelt- eller trippelbindinger mellom karbonene).
Blant de mest fremtredende fosfoglyseridene i cellemembraner har vi: sfingomyelin, fosfatidylinositol, fosfatidylserin og fosfatidyletanolamin.
Matvarer som er rike på disse biologiske molekylene, er hvitt kjøtt som blant annet fisk, eggeplommer, noe organkjøtt, skalldyr og nøtter..
Artikkelindeks
Et fosfoglyserid består av fire grunnleggende strukturelle elementer. Førstnevnte er et skjelett som fettsyrer, et fosfat og en alkohol er bundet til - det siste er bundet til fosfat.
Fosfoglyserid-ryggraden kan dannes fra glyserol eller sfingosin. Den første er en alkohol med tre karbon, mens den andre er en annen alkohol med en mer kompleks struktur.
I glyserol forestres hydroksylgruppene ved karbon 1 og 2 av to store kjedekarboksylgrupper av fettsyrer. Det manglende karbonet, plassert i posisjon 3, forestres med en fosforsyre.
Selv om glyserol ikke har et asymmetrisk karbon, er alfakarbonene ikke stereokjemisk identiske. Dermed gir forestring av et fosfat på det tilsvarende karbon asymmetri til molekylet.
Fettsyrer er molekyler sammensatt av hydrokarbonkjeder med varierende lengde og grad av umettethet, og ender i en karboksylgruppe. Disse egenskapene varierer betydelig og bestemmer egenskapene..
Fettsyrekjeden er lineær hvis den er av mettet type eller hvis den er umettet i stilling trans. I kontrast tilstedeværelsen av en dobbeltbinding av typen cis skaper en vri i kjeden, slik at den ikke lenger trenger å bli representert lineært, som ofte er tilfelle.
Fettsyrer med dobbelt- eller trippelbindinger påvirker tilstanden og de fysisk-kjemiske egenskapene til biologiske membraner betydelig.
Hvert av de nevnte elementene er forskjellige i deres hydrofobe egenskaper. Fettsyrer, som er lipider, er hydrofobe eller apolare, noe som betyr at de ikke blandes med vann.
I kontrast tillater resten av elementene i fosfolipider dem å samhandle i et miljø, takket være deres polære eller hydrofile egenskaper..
På denne måten klassifiseres fosfoglyserider som amfipatiske molekyler, noe som betyr at den ene enden er polær og den andre er apolar..
Vi kan bruke analogien til en kamp eller en kamp. Kampens hode representerer det polare hodet som består av det ladede fosfatet og erstatningene av det på fosfatgruppen. Forlengelsen av fyrstikken er representert av en ikke-polær hale, dannet av hydrokarbonkjeder.
Grupper av polar art er ladet til pH 7, med en negativ ladning. Dette skyldes ioniseringsfenomenet til fosfatgruppen, som har en pk nær 2, og anklagene til de forestrede gruppene. Antall ladninger avhenger av typen fosfoglyserid som er undersøkt.
Lipider er hydrofobe biomolekyler som presenterer løselighet i organiske løsningsmidler - for eksempel kloroform..
Disse molekylene har en rekke funksjoner: spille en rolle som drivstoff ved å lagre konsentrert energi; som signalmolekyler; og som strukturelle komponenter i biologiske membraner.
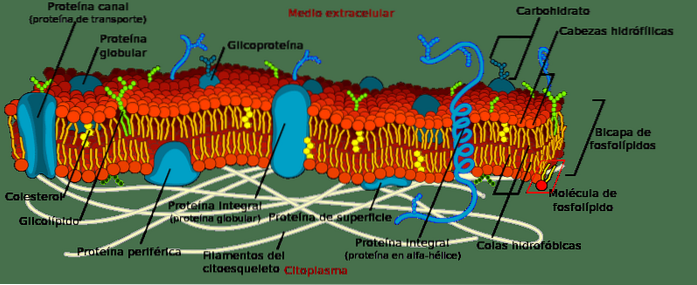
I naturen er den mest utbredte gruppen lipider fosfoglyserider. Deres hovedfunksjon er strukturell, siden de er en del av alle cellemembraner.
Biologiske membraner er gruppert i form av et dobbeltlag. Noe som betyr at lipider er gruppert i to lag, hvor deres hydrofobe haler vender mot det indre av dobbeltlaget og de polære hodene vender mot det ytre og indre av cellen..
Disse strukturene er avgjørende. De avgrenser cellen og er ansvarlig for utveksling av stoffer med andre celler og med det ekstracellulære miljøet. Imidlertid inneholder membranene andre lipidmolekyler enn fosfoglyserider, og også molekyler av protein-karakter som formidler aktiv og passiv transport av stoffer..
I tillegg til å være en del av biologiske membraner, er fosfoglyserider assosiert med andre funksjoner i mobilmiljøet. Noen veldig spesifikke lipider er en del av myelinens membraner, stoffet som dekker nervene.
Noen kan fungere som meldinger i fangst og overføring av signaler til mobilmiljøet.
Syntesen av fosfoglyserider utføres med utgangspunkt i mellomliggende metabolitter så som fosfatidinsyremolekylet og også triacylglyseroler..
Et aktivert nukleotid CTP (cytidintrifosfat) danner et mellomprodukt kalt CDP-diacylglyserol, der pyrofosfatreaksjonen favoriserer reaksjonen til høyre.
Delen kalt fosfatidyl reagerer med visse alkoholer. Produktet av denne reaksjonen er fosfoglyserider, inkludert fosfatidylserin eller fosfatidylinositol. Fosfatidyletanolamin eller fosfatidylkolin kan fås fra fosfatidylserin.
Imidlertid er det alternative veier for å syntetisere sistnevnte fosfoglyserider. Denne veien involverer aktivering av kolin eller etanolamin ved binding til CTP..
Deretter oppstår en reaksjon som forener dem med fosfatidat og oppnår fosfatidyletanolamin eller fosfatidylkolin som sluttprodukt..
Nedbrytningen av fosfoglyserider utføres av enzymer som kalles fosfolipaser. Reaksjonen innebærer frigjøring av fettsyrene som utgjør fosfoglyserider. I alle vev av levende organismer foregår denne reaksjonen konstant.
Det er flere typer fosfolipaser, og de er klassifisert i henhold til fettsyren som de frigjøres. Etter dette klassifiseringssystemet skiller vi mellom lipaser A1, A2, C og D..
Fosfolipaser er allestedsnærværende i naturen, og vi finner dem i forskjellige biologiske enheter. Tarmsaft, sekresjoner av visse bakterier og slangegift er eksempler på stoffer med høye fosfolipaser..
Sluttproduktet av disse nedbrytningsreaksjonene er glyserol-3-fosfat. Dermed kan disse frigjorte produktene pluss frie fettsyrer gjenbrukes for å syntetisere nye fosfolipider eller ledes til andre metabolske veier..
Forbindelsen beskrevet ovenfor er det enkleste fosfoglyseridet og kalles fosfatidat, eller også diacylglyserol-3-fosfat. Selv om det ikke er veldig rikelig i det fysiologiske miljøet, er det et nøkkelelement for syntesen av mer komplekse molekyler.
Fra det enkleste fosfoglyseridmolekylet kan biosyntese av mer komplekse elementer oppstå, med svært viktige biologiske roller..
Fosfatgruppen av fosfatidat forestres med hydroksylgruppen av alkoholer - den kan være en eller flere. De vanligste alkoholene til folfoglyseridene er serin, etanolamin, kolin, glyserol og inositol. Disse derivatene vil bli beskrevet nedenfor:
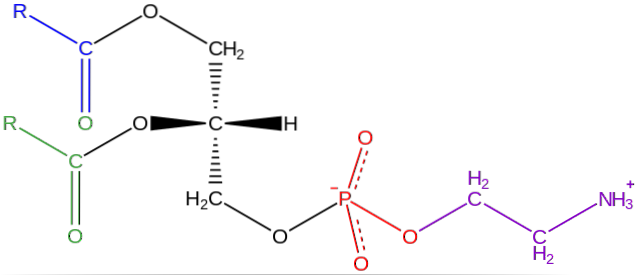
I cellemembraner som er en del av humant vev, er fosfatidyletanolamin en fremtredende komponent i disse strukturene.
Den består av en alkohol forestret av fettsyrer i hydroksylene i posisjon 1 og 2, mens vi i posisjon 3 finner en fosfatgruppe forestret med aminoalkoholet etanolamin.
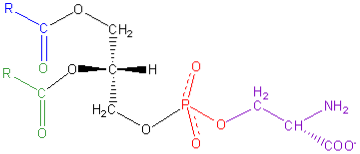
Generelt finnes dette fosfoglyseridet i monolaget som vender mot det indre - det vil si den cytosoliske siden - av cellene. Under prosessen med programmert celledød, endres fordelingen av fosfatidylserin og finnes over hele celleoverflaten.
Fosfatidylinositol er et fosfolipid som finnes i lave proporsjoner både i cellemembranen og i membranene til de subcellulære komponentene. Det har vist seg å delta i mobilkommunikasjonshendelser og forårsake endringer i det indre miljøet i cellen.

I gruppen fosfolipider er sfingomyelin det eneste fosfolipidet som er tilstede i membraner hvis struktur ikke er avledet fra alkoholglyserolen. I stedet består skjelettet av sfingosin.
Strukturelt tilhører denne siste forbindelsen gruppen aminoalkoholer og har en lang karbonkjede med dobbeltbindinger..
I dette molekylet er aminogruppen på ryggraden festet til en fettsyre via en amidlignende binding. Til sammen forestres den primære hydroksylgruppen i skjelettet til fosfotylkolin.
Plasmallogener er fosfoglyserider med hoder dannet hovedsakelig av etanolamin, kolin og serin. Funksjonene til disse molekylene er ikke fullstendig belyst, og det er lite informasjon om dem i litteraturen..
Fordi vinylethergruppen lett oksyderes, er plasmalogen i stand til å reagere med oksygenfrie radikaler. Disse stoffene er produkter med gjennomsnittlig cellulær metabolisme og har vist seg å skade cellulære komponenter. I tillegg har de også vært relatert til aldringsprosesser.
Derfor er en mulig funksjon av plasmalogener å fange frie radikaler som potensielt kan ha negative effekter på celleintegritet..



Ingen har kommentert denne artikkelen ennå.