De steroidhormoner De er stoffer laget av kjertler med intern sekresjon og som dumpes direkte i sirkulasjonsstrømmen, som fører dem til vevet der de utøver sine fysiologiske effekter. Det generiske navnet stammer fra det faktum at det har en steroidkjerne i sin grunnleggende struktur..
Kolesterol er forløperstoffet som alle steroidhormoner syntetiseres fra, som er gruppert i gestagener (for eksempel progesteron), østrogener (estron), androgener (testosteron), glukokortikoider (kortisol), mineralokortikoider (aldosteron) og vitamin D.
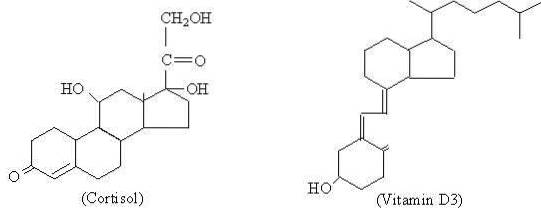
Selv om de forskjellige steroidhormonene presenterer molekylære forskjeller mellom dem, som er det som gir dem forskjellige funksjonelle egenskaper, kan det sies at de har en grunnleggende struktur som er felles for dem, og som er representert av cyklopentaneperhydrofenanthren med 17 karbonatomer..
Artikkelindeks
Steroider er organiske forbindelser av svært mangfoldig natur som har det som kan betraktes som en kjernekjerne som består av fusjonen av tre ringer med seks karbonatomer (cykloheksaner) og ett av fem karbonatomer (cyklopentan)..
Denne strukturen er også kjent som "cyclopentaneperhydrophenanthrene". Ettersom ringene er gjensidig forbundet, er det totale antallet karbonatomer som utgjør det 17; Imidlertid har de fleste naturlige steroider metylgrupper ved karbon 13 og 10, som representerer henholdsvis karbon 18 og 19..

Mange av de naturlige steroidforbindelsene har også en eller flere grupper med alkoholfunksjon i ringstrukturen og kalles derfor steroler. Blant dem er kolesterol, som har en alkoholfunksjon ved karbon 3 og en hydrokarbonsidekjede med 8 karbonatomer festet til karbon 17; atomer som er nummerert fra 20 til 27.
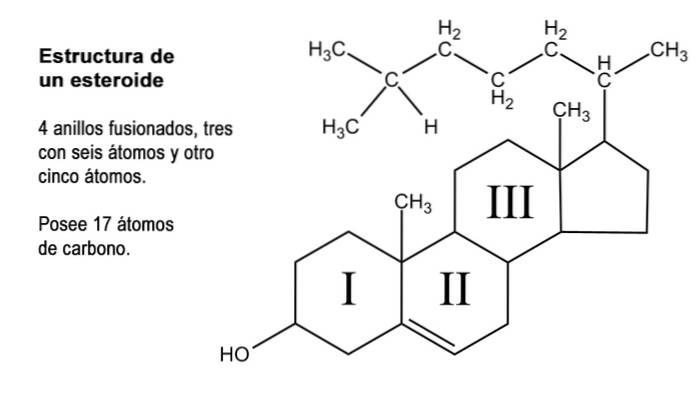
I tillegg til disse 17 karbonene, kan steroidhormoner ha 1, 2 eller 4 flere av disse atomer i strukturen, for hvilke tre typer steroider er gjenkjent, nemlig: C21, C19 og C18.
C21s, som progesteron og binyrebarksteroider (glukokortikoider og mineralokortikoider), er avledet fra "gravidan". Den har 21 karbonatomer fordi til 17 av den basiske ringen er tilsatt to av metylgruppene i karbon 13 og 10, og to karbonatomer i sidekjeden festet til C17 som opprinnelig, i kolesterol, var 8 karbonatomer.
C19-ene tilsvarer kjønnshormoner med androgen aktivitet og er avledet fra "androstan" (19 karbonatomer), som er strukturen som gjenstår når gravidan mister de to karbonene i C17-sidekjeden, som erstattes av en hydroksyl- eller en ketongruppe.
C18-steroider er kvinnelige hormoner eller østrogener som hovedsakelig syntetiseres i kvinnelige kjønnsorganer, og hvis fremragende karakteristikk, med hensyn til de to andre typene steroider, er fraværet av metylen som er tilstede i sistnevnte festet til karbonet i posisjon 10.
Under syntesen fra kolesterol produseres enzymatiske modifikasjoner som endrer antall karbonatomer og fremmer dehydrogeneringer og hydroksyleringer av spesifikke karboner i strukturen..
Cellene som produserer steroidhormoner ligger hovedsakelig i cortex i binyrene, hvor glukokortikoider som kortisol, mineralokortikoider som aldosteron og mannlige kjønnshormoner som dehydroepiandrosteron og androstenedion produseres..
Mannlige kjønnskirtler er ansvarlige for produksjonen av androgener, inkludert de nevnte hormonene og testosteron, mens de modne ovariefolliklene produserer progesteron og østrogener..
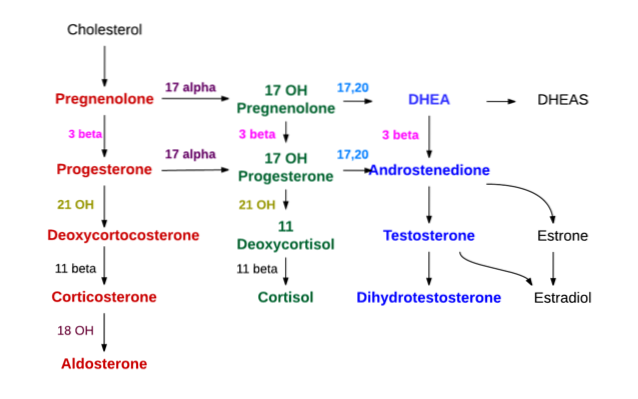
Syntesen av alle steroidhormoner starter fra kolesterol. Dette molekylet kan syntetiseres av celler som produserer steroidhormoner, men for det meste oppnås det av disse cellene fra lavdensitetslipoproteiner (LDL) som er tilstede i sirkulerende plasma..
Tre lag skiller seg ut i binyrebarken, kjent fra utsiden som henholdsvis glomerulær, fascikulær og retikulær son..
I glomerulæren syntetiseres mineralokortikoider (aldosteron) hovedsakelig, i fascikulære glukokortikoider som kortikosteron og kortisol, og i retikulære androgener som dehydroepiandrosteron og androstenedion.
Det første trinnet i syntesen skjer i mitokondriene og består av virkningen av et enzym kalt kolesterol desmolase, som tilhører cytokrom P450 superfamilien og også kjent som "P450scc" eller "CYP11A1", som fremmer eliminering av 6 av karbonatomer av sidekjeden festet til C17.
Med virkningen av desmolase omdannes kolesterol (med 27 karbonatomer) til pregnenolon, som er en forbindelse med 21 karbonatomer og representerer den første av C21-steroider.
Pregnenolon beveger seg til det glatte endoplasmatiske retikulumet, der ved virkning av enzymet 3β-hydroksysteroid dehydrogenase gjennomgår dehydrogenering i hydroksylen av alkoholgruppen av karbon 3, og blir progesteron.
På grunn av virkningen av 21β-hydroksylase, også kalt “P450C21” eller “CYP21A2”, hydroksyleres progesteron ved karbon 21 og transformeres til 11-deoksykortikosteron, som går tilbake til mitokondriene, og til hvilket enzymet 11β-hydroksylase (“P450C11 ”Eller” CYP11B1 ”) konverterer til kortikosteron.
En annen syntese i den fascikulære sonen som ikke ender i kortikosteron, men i kortisol, oppstår når gravidenolon eller progesteron hydroksyleres i posisjon 17 av 17a-hydroksylase ("P450C17" eller "CYP17") og omdannes 17-hydroksypregnolon eller 17-hydroksyprogesteron.
Det samme enzymet som allerede er nevnt, 3β-hydroksysteroid dehydrogenase, som omdanner pregnenolon til progesteron, konverterer også 17-hydroksypregnolon til 17-hydroksyprogesteron.
Sistnevnte bæres suksessivt av de to siste enzymene i banen som produserer kortikosteron (21β-hydroksylase og 11β-hydroksylase) til henholdsvis deoksykortisol og kortisol..
De viktigste glukokortikoider produsert i zona fascicular av binyrebarken er kortikosteron og kortisol. Begge stoffene, men spesielt kortisol, viser et bredt spekter av handlinger som påvirker metabolisme, blod, forsvar og sårhelingsresponser, beinmineralisering, fordøyelseskanalen, sirkulasjonssystemet og lungene..
Når det gjelder metabolisme, stimulerer kortisol lipolyse og frigjøring av fettsyrer som kan brukes i leveren til dannelse av ketonlegemer og LDL-proteiner; reduserer glukoseopptak og lipogenese i fettvev og glukoseopptak og bruk i muskler.
Det fremmer også proteinkatabolisme i periferien: i bindevev, muskel og beinmatrise, og frigjør derved aminosyrer som kan brukes i leveren for syntese av plasmaproteiner og for glukoneogenese. Det stimulerer dessuten absorpsjon av tarmsukker ved å øke produksjonen av SGLT1-transportører..
Akselerert tarmsukkerabsorpsjon, økt leverproduksjon og redusert utnyttelse av dette karbohydratet i muskel- og fettvev favoriserer en økning i plasmaglukosenivået..
Når det gjelder blodet, favoriserer kortisol koaguleringsprosessen, stimulerer dannelsen av nøytrofile granulocytter og hemmer eosinofiler, basofiler, monocytter og T-lymfocytter. Det hemmer også frigjøring av inflammatoriske mediatorer som prostaglandiner, interleukiner, lymfokiner, histamin og serotonin.
Generelt kan det sies at glukokortikoider forstyrrer immunresponsen, av hvilken grunn de kan brukes terapeutisk i de tilfeller der denne responsen er overdrevet eller upassende, for eksempel i tilfelle av autoimmune sykdommer eller i organtransplantasjoner for å redusere avvisning.
Androgensyntese på nivået av binyrebarken forekommer hovedsakelig på nivået av retikulær sone og fra 17-hydroksypregnolon og 17-hydroksyprogesteron.
Det samme 17α-hydroksylaseenzymet, som produserer de to nevnte stoffene, har også 17,20 lyase-aktivitet, som fjerner de to karbonene i C17-sidekjeden og erstatter dem med en ketogruppe (= O).
Denne siste handlingen reduserer karbonantallet med to og produserer C19-steroider. Hvis handlingen er på 17-hydroksypregnolon, er resultatet dehydroepiandrosteron; hvis det berørte stoffet tvert imot er hydroksyprogesteron, vil produktet være androstenion.
Begge forbindelsene er en del av de såkalte 17-ketosteroider, da de har en ketongruppe ved karbon 17.
3β-hydroksysteroiddehydrogenase konverterer også dehydroepiandrosteron til androstenedion, men det vanligste er at førstnevnte omdannes til dehydroepiandrosteronsulfat av en sulfokinase, tilstede nesten utelukkende i retikulær sone.
Zona glomerularis mangler 17a-hydroksylaseenzymet, og kan ikke syntetisere 17-hydroksysteroidforløperne til kortisol og kjønnshormoner. Det har heller ikke 11β-hydroksylase, men det har et enzym som kalles aldosteronsyntetase som sekvensielt kan produsere kortikosteron, 18-hydroksykortikosteron og mineralokortikoid aldosteron..
Det viktigste mineralokortikoid er aldosteron syntetisert i zona glomerularis i binyrebarken, men glukokortikoider viser også mineralokortikoid aktivitet.
Aldosteronets mineralokortikoidaktivitet utvikler seg på nivået av det rørformede epitelet i den distale nefronen, hvor det fremmer sekresjon av natriumreabsorpsjon (Na +) og kalium (K +), og bidrar dermed til bevaring av nivåene av disse ionene i kroppsvæskene..
Testikulær androgensyntese skjer på nivå med Leydig-celler. Testosteron er det viktigste androgenhormonet som produseres i testiklene. Dens syntese involverer den første produksjonen av androstenedion som tidligere beskrevet for syntesen av androgener på nivået av binyrebarken..
Androstenedion omdannes til testosteron ved virkningen av enzymet 17β-hydroksysteroiddehydrogenase, som erstatter ketongruppen av karbon 17 med en hydroksylgruppe (OH).
I noen vev som tjener som et mål for testosteron, reduseres det av en 5a-reduktase til dihydrotestosteron, med større androgen kraft.
Denne syntesen skjer syklisk med endringene som skjer under den kvinnelige seksuelle syklusen. Syntesen skjer i follikkelen som i løpet av hver syklus modnes for å frigjøre et egg og deretter produsere det tilsvarende corpus luteum.
Østrogener syntetiseres i de granulære cellene i den modne follikkelen. Den modne follikelen har celler i theca som produserer androgener som androstenedion og testosteron..
Disse hormonene diffunderer i nærliggende granulosaceller, som har aromataseenzymet som omdanner dem til estron (E1) og 17β-østradiol (E2). Fra begge er estriol syntetisert.
Androgener og østrogener har som hovedfunksjon utviklingen av henholdsvis mannlige og kvinnelige seksuelle egenskaper. Androgener har anabole effekter som fremmer syntesen av strukturelle proteiner, mens østrogener favoriserer ossifikasjonsprosessen.
Østrogenene og progesteronet som frigjøres under den kvinnelige seksuelle syklusen, er ment å forberede kvinnens kropp for en eventuell graviditet som et resultat av befruktningen av det modne egget som frigjøres under eggløsningen..
Hvis du trenger å oppdatere hukommelsen om hormonens virkningsmekanisme, anbefales det å se følgende video før du fortsetter å lese.
Virkningsmekanismen til steroidhormoner er ganske lik i dem alle. Når det gjelder lipofile forbindelser, oppløses de uten problemer i lipidmembranen og trenger inn i cytoplasmaet i målcellene, som har spesifikke cytoplasmatiske reseptorer for hormonet som de må reagere på..
Når hormonreseptorkomplekset er dannet, krysser det kjernemembranen og binder seg i genomet, på samme måte som en transkripsjonsfaktor, med et hormonresponselement (HRE) eller primært responsgen, som igjen Det kan noen ganger regulere andre gener kalt sekundærrespons.
Sluttresultatet er markedsføring av transkripsjon og syntese av messenger-RNA som blir oversatt i ribosomene i det grove endoplasmatiske retikulumet som ender med å syntetisere proteinene indusert av hormonet..
Aldosteronets virkning utøves hovedsakelig på nivået av den siste delen av det distale røret og i samlekanalene, der hormonet fremmer Na + reabsorpsjon og K-sekresjon+.
I luminalmembranen til de viktigste rørformede cellene i denne regionen er det epiteliale Na + -kanaler og K + -kanaler av "ROMK" -typen. Nyre ytre medullær kaliumkanal).
Den basolaterale membranen har Na + / K + ATPase-pumper som kontinuerlig trekker Na + fra cellen inn i det basolaterale interstitielle rommet og introduserer K + i cellen. Denne aktiviteten holder den intracellulære konsentrasjonen av Na + veldig lav og favoriserer dannelsen av en konsentrasjonsgradient for dette ionet mellom lumen i tubuli og cellen..
Denne gradienten tillater Na + å bevege seg mot cellen gjennom epitelkanalen, og siden Na + passerer alene, forblir det for hvert ion som beveger seg en ukompensert negativ ladning som får tubulens lumen til å bli negativ med hensyn til interstitium. Det vil si at en transepitelial potensialforskjell skapes med det negative lyset.
Denne negativiteten til lyset favoriserer utgangen av K + som beveger seg av dens høyere konsentrasjon i cellen, og negativiteten til lyset utskilles mot tubulens lumen for å endelig utskilles. Det er denne Na + reabsorpsjonen og K + sekresjonsaktiviteten som reguleres av virkningen av aldosteron..
Aldosteron som er tilstede i blodet og frigjøres fra zona glomerularis som respons på virkningen av angiotensin II, eller til hyperkalemi, trenger inn i hovedcellene og binder seg med sin intracytoplasmiske reseptor.
Dette komplekset når kjernen og fremmer transkripsjon av gener hvis ekspresjon vil ende opp med å øke syntesen og aktiviteten til Na + / K + pumper, epiteliale Na + kanaler og ROMK K + kanaler, så vel som andre proteiner. Respons som vil ha den generelle effekten av retensjonen av Na + i kroppen og økningen i K-utskillelse i urinen+.



Ingen har kommentert denne artikkelen ennå.