De prosessminne eller instrumental er den som lagrer prosedyrer, ferdigheter eller motoriske eller kognitive ferdigheter som lar folk samhandle med miljøet.
Det er en type ubevisst langtidsminne, og gjenspeiler måten å gjøre ting på (motoriske ferdigheter). For eksempel: å skrive, sykle, kjøre bil, spille et instrument, blant andre.

Minnesystemer er vanligvis delt inn i to typer: deklarativt minne og ikke-deklarativt eller implisitt minne. Den første er en som lagrer informasjon som kan kommuniseres muntlig, bestående av bevisst læring.
På den annen side er den andre typen et minne som er vanskelig å verbalisere eller forvandle til bilder. Innenfor det er prosessminne. Dette aktiveres når en oppgave må utføres, og de lærte funksjonene er vanligvis ferdigheter som er automatiserte.
Hovedhjernesubstratet til prosessuelt minne er striatum, basalganglier, premotorisk cortex og lillehjernen..
Utviklingen av prosedyreminne skjer i større grad i barndommen. Og det blir kontinuerlig modifisert av daglige erfaringer og praksis. Det er sant at det i voksen alder er vanskeligere å tilegne seg denne typen ferdigheter enn i barndommen, siden det krever en ekstra innsats.
Artikkelindeks
Prosedyreminne består av vaner, ferdigheter og motoriske ferdigheter som motorsystemet tilegner seg og inkorporerer i sine egne kretser. For at denne typen minne skal tilegnes, er det nødvendig å gi flere treningsforsøk som gjør det mulig å automatisere ferdighetene..
Kunnskap utvikler seg ubevisst og moduleres kontinuerlig av erfaring. Dermed tilpasser de seg gjennom hele livet til gjentatt praksis..
I mer avanserte stadier gjør praksis kognitive eller motoriske ferdigheter mer presise og raskere. Dette blir en vane, en oppførsel som kjører automatisk.
Det ser ut til å være to typer prosedyreminne, med forskjellige hovedsteder i hjernen..
Den første refererer til anskaffelse av vaner og ferdigheter. Evnen til å utvikle stereotype atferdsmessige repertoarer som å skrive, lage mat, spille piano ... Denne typen prosedyreminne handler om målrettet atferd, og ligger i hjernens striated system..
Det andre er et mye enklere system. Det refererer til spesifikke sensorimotoriske tilpasninger, det vil si å justere refleksene våre eller utvikle kondisjonerte reflekser.
Dette er kroppsjusteringer som tillater utførelse av fine og presise bevegelser, samt kondisjonerte reflekser. Den ligger i cerebellar-systemet.
Prosedyreminne begynner å dannes tidlig når du lærer å gå, snakke eller spise. Slike ferdigheter blir gjentatt og inngrodd på en slik måte at de gjøres automatisk. Det er ikke nødvendig å bevisst tenke på hvordan man skal utføre slike motoriske aktiviteter.
Det er vanskelig å si når du lærte å utføre denne typen handlinger. De læres vanligvis i tidlig barndom og fortsetter å bli utført ubevisst.
Tilegne seg disse ferdighetene krever trening, selv om det er sant at opplæring ikke alltid sikrer at ferdighetene blir utviklet. Vi kan si at prosessuell læring er anskaffet når atferd endres takket være trening.
Tilsynelatende er det strukturer i hjernen vår som styrer den første læringen av prosessuelle minner, deres sene læring og deres automatisering.
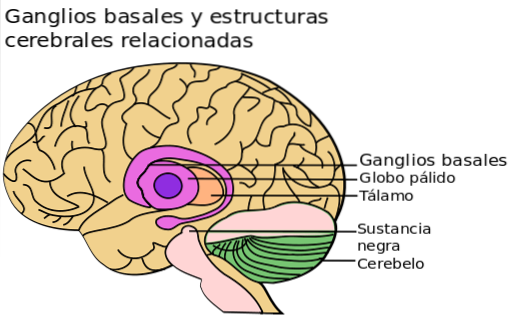
Når vi lærer en vane, aktiveres et område av hjernen vår som kalles basalganglier. Basalganglier er subkortikale strukturer som har flere forbindelser til hele hjernen..
Spesielt tillater de utveksling av informasjon mellom nedre hjerneområder (som hjernestammen) og høyere områder (for eksempel hjernebarken).
Denne strukturen ser ut til å spille en selektiv rolle i prosessuell læring av vaner og ferdigheter. Den deltar også i andre ikke-deklarative minnesystemer, for eksempel klassisk eller operant kondisjonering.
Innenfor basalganglier skiller en region som kalles den striated nucleus seg ut i tilegnelsen av vaner. Den mottar informasjon fra det meste av hjernebarken, så vel som andre deler av basalganglier..
Striatum er delt inn i assosiativ striatum og sensorimotorisk striatum. Begge har forskjellige funksjoner innen læring og automatisering av ferdigheter.
Når vi er i de tidlige stadiene av prosessuell læring, blir den assosiative striatum aktivert. Interessant, da aktiviteten er trening og læring, reduserer dette området sin aktivitet. Dermed, når vi lærer å kjøre, blir den assosiative striatum aktivert.
For eksempel i en studie av Miyachi et al. (2002), ble det funnet at hvis det associative striatumet ble midlertidig inaktivert, kunne ikke nye bevegelsessekvenser læres. Emnene kunne imidlertid utføre allerede innlærte motoriske mønstre.
I senere stadier av prosessuell læring blir en annen struktur aktivert: sensorimotor striatum. Dette området har et aktivitetsmønster motsatt det assosiative striatum, det vil si at det aktiveres når ferdigheten allerede er anskaffet og er automatisk..
Når denne evnen til å kjøre har blitt tilstrekkelig opplært og allerede er automatisk, reduserer den assosiative striatum sin aktivitet mens aktiveringen av sensorimotorisk striatum øker..
I tillegg er det funnet at en midlertidig blokkering av sensorimotor striatum forhindrer utførelsen av de lærte sekvensene. Selv om det ikke forstyrrer læringen av nye ferdigheter.
Imidlertid ser det ut til å være ett skritt til. Det er observert at når en oppgave allerede er veldig godt lært og automatisert, slutter også neuronene i sensorimotor striatum å svare.
Hva skjer da? Tilsynelatende, når en oppførsel er veldig godt lært, blir hjernebarken (cortex) for det meste aktivert. Mer spesifikt motor- og premotorområdet.
Selv om dette også ser ut til å avhenge av hvor kompleks bevegelsessekvensen er. Dermed, hvis bevegelsene er enkle, blir hjernebarken overveiende aktivert.
På den annen side, hvis sekvensen er veldig kompleks, fortsetter noen nevroner i sensorimotor striatum å bli aktivert. I tillegg til å aktivere motor- og premotoriske regioner i hjernebarken som støtte.
På den annen side har det vist seg at det er en reduksjon i aktiviteten til hjerneområdene som styrer oppmerksomheten (prefrontal og parietal) når vi utfører høys automatiserte oppgaver. Mens, som nevnt, øker aktiviteten i motor- og førmotoriske områder.
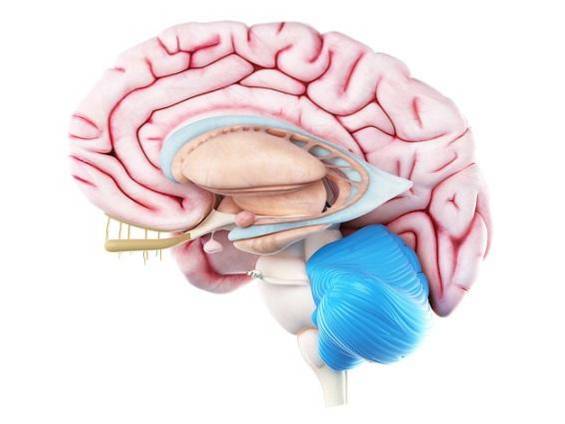
Cerebellum ser også ut til å delta i prosedyreminne. Spesielt deltar den ved å foredle og gjøre de lærte bevegelsene mer presise. Det vil si at det gir oss mer smidighet når vi utfører motoriske ferdigheter.
I tillegg hjelper det å lære nye motoriske ferdigheter og konsolidere dem gjennom Purkinje-celler.
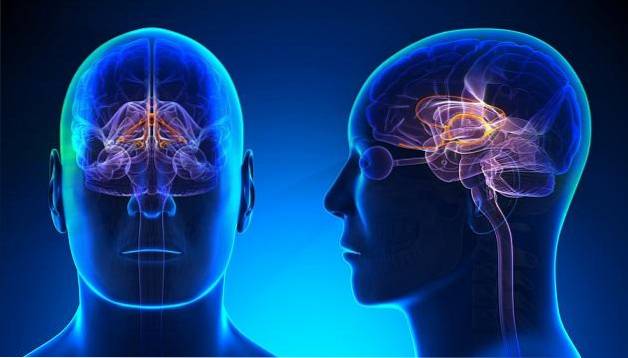
Som i andre minnesystemer spiller det limbiske systemet en viktig rolle i prosessuell læring. Dette er fordi det er relatert til prosesser med motivasjon og følelser..
Av denne grunn, når vi er motivert eller interessert i å lære en oppgave, lærer vi den lettere, og den blir liggende lenger i minnet..
Det er vist at når vi tilegner oss læring, endres forbindelsene og strukturene til nevronene.
På denne måten, gjennom en rekke prosesser, begynner de lærte ferdighetene å bli en del av langtidsminnet, reflektert i en omorganisering av nevrale kretser..
Visse synapser (forbindelser mellom nevroner) styrkes og andre svekkes, samtidig som de dendrittiske pigger av nevroner endrer seg i størrelse, forlenger.
På den annen side er tilstedeværelsen av dopamin viktig for prosessminnet. Dopamin er en nevrotransmitter i nervesystemet som har flere funksjoner, inkludert økende motivasjon og følelser av belønning. I tillegg til å tillate bevegelse, og selvfølgelig læring.
Det letter hovedsakelig læringen som skjer takket være belønninger, for eksempel å lære å trykke på en bestemt knapp for å få mat.
Det er forskjellige tester for å evaluere prosessuell minnekapasitet hos mennesker. Studier bruker ofte slike tester som sammenligner ytelse mellom pasienter med hukommelsesproblemer og friske mennesker.
De mest brukte oppgavene for å evaluere prosessminne er:
I denne oppgaven måles prosessuell kognitiv læring. Deltakeren får presentert fire forskjellige korttyper der forskjellige geometriske figurer vises. Hvert kort representerer en viss sannsynlighet for at det regner eller skinner.
I neste trinn presenteres emnet med tre grupperte kort. Dette må finne ut om det er mer sannsynlig at det vil være sol eller regn når vi tar dataene sammen.
Etter svaret ditt vil sensoren fortelle deg om svaret var riktig eller ikke. Derfor lærer deltakeren i hvert forsøk gradvis å identifisere hvilke kort som er assosiert med større sannsynlighet for sol eller regn..
Pasienter med forandringer i basale ganglier, slik som de med Parkinsons sykdom, klarer ikke gradvis å lære denne oppgaven, selv om deres eksplisitte minne er intakt.
Denne oppgaven vurderer læring av sekvenser. I den presenteres visuelle stimuli på en skjerm, vanligvis bokstaver (ABCD ...) Deltakeren får beskjed om å se på posisjonen til en av dem (for eksempel B).
Deltakeren må trykke en av fire taster, avhengig av hvor målstimulansen er, så raskt som mulig. Venstre mellom- og pekefinger, og høyre pekefinger og langfinger brukes.
Først er posisjonene tilfeldige, men i neste fase følger de et bestemt mønster. For eksempel: DBCACBDCBA ... Så, etter flere studier, bør pasienten lære de nødvendige bevegelsene og automatisere dem.
Denne oppgaven utføres med en spesiell enhet som har en roterende plate. I den ene delen av platen er det et metallspiss. Deltakeren må plassere en stang i metallspissen så lenge som mulig, uten å glemme at platen utfører sirkulære bevegelser som må følges.
God hånd-øye-koordinering er nødvendig i denne oppgaven. Det tester evnen til å lære en bestemt motorisk ferdighet, for eksempel å spore omrisset til en stjerne. Imidlertid, for denne oppgaven kan deltakeren bare se refleksjonen av bildet han tegner i et speil.
Feil er vanlig i begynnelsen, men etter flere repetisjoner kontrolleres bevegelser ved å observere egen hånd og tegningen i speilet. Hos friske pasienter blir det gjort færre og færre feil.

Det har blitt demonstrert mye at prosessminnet konsolideres gjennom en offline prosess. Det vil si at vi fikser instrumentelle minner i hvileperioder mellom motorisk trening, spesielt under søvn..
På denne måten er det observert at motoroppgaver ser ut til å forbedre seg markant når de blir evaluert etter et hvileintervall.
Dette skjer med alle typer minne. Etter en periode med praksis har det blitt funnet gunstig å hvile, slik at det du har lært tar tak. Disse effektene forbedres ved å hvile like etter treningsperioden.
Prosedyreminne har komplekse forhold til bevissthet. Vi refererer tradisjonelt til denne typen minne som et ubevisst minne som ikke involverer innsats..
Imidlertid har eksperimentelle studier vist at nevral aktivering skjer før bevisst planlegging av bevegelsen som skal utføres oppstår.
Det vil si at det bevisste ønsket om å utføre en bevegelse faktisk er en "illusjon". I følge forskjellige studier kan faktisk det å være "bevisst" på våre automatiske bevegelser faktisk påvirke utførelsen av oppgaven negativt.
På denne måten, når vi blir klar over vår bevegelsesrekkefølge, forverres vi noen ganger i ytelse og gjør flere feil. Av denne grunn understreker mange forfattere fremfor alt at prosedyreminne, når det allerede er godt etablert, ikke krever oppmerksomhet eller tilsyn med handlingene selv for å gjøre det godt..
Det er et sett med både kortikale og subkortikale strukturer som griper inn i forskjellige funksjoner i prosessminnet. En selektiv lesjon av noen av dem gir forskjellige forstyrrelser i motoriske funksjoner, som lammelse, apaksi, ataksi, skjelving, koreiske bevegelser eller dystoni.
Mange studier har analysert patologiene som påvirker hukommelsen med det formål å vite hvilke typer eksisterende minner og hvordan de fungerer.
I dette tilfellet er de mulige konsekvensene som en funksjonsfeil i basalganglier eller andre strukturer kan ha på læring og utføring av oppgaver blitt undersøkt..
For dette brukes forskjellige evalueringstester i de forskjellige studiene som sammenligner friske mennesker og andre med noe svekkelse av prosessminnet. Eller pasienter med prosedyreminnehemming og andre pasienter med nedsatt hukommelse.
For eksempel er det i Parkinsons sykdom et underskudd av dopamin i striatum og abnormiteter i utførelsen av visse minneoppgaver er observert. Problemer kan også oppstå i Huntingtons sykdom, der det er skade på forbindelsene mellom basalganglier og hjernebarken..
Det vil også oppstå vanskeligheter hos pasienter med hjerneskade på noen av hjernestrukturene som er involvert (for eksempel som produseres av hjerneslag).
Imidlertid er den eksakte rollen til basalganglier i læringsbevegelsen i dag noe kontroversiell..
Det er funnet at visse hjerneområder under motorisk læring aktiveres hos friske deltakere. Noen av dem var dorsolateral prefrontal cortex, det supplerende motorområdet, den fremre cingulate cortex ... samt basalganglier..
Imidlertid ble andre forskjellige områder aktivert hos Parkinsons pasienter (som lillehjernen). I tillegg var striatum og basalganglier inaktive. Det ser ut til at kompensasjon oppstår gjennom kortico-cerebellar-systemet, siden kortico-striatal-banen er skadet.
Hos pasienter med denne sykdommen og med Huntingtons sykdom er det også observert en større aktivering av hippocampus og thalamisk-kortikale baner..
I en annen studie vurderte de pasienter som hadde fått hjerneslag som involverte basalganglier og sammenlignet dem med friske deltakere.
De fant ut at berørte pasienter lærer motoriske sekvenser saktere, det tar lengre tid å gi svar, og svarene er mindre nøyaktige enn for friske deltakere..
Tilsynelatende er forklaringene gitt av forfatterne at disse individene har problemer med å dele motorsekvensen i organiserte og koordinerte elementer. Dermed er svarene deres uorganiserte og det tar lenger tid å utdype dem..


Ingen har kommentert denne artikkelen ennå.