De plastokinon (PQ) er et organisk lipidmolekyl, spesielt et isoprenoid fra kinonfamilien. Faktisk er det et sidekjede flerumettet derivat av kinon som deltar i fotosystem II av fotosyntese..
Ligger i kloroplasterens tylakoidmembran, har den apolar karakter og er veldig aktiv på molekylært nivå. Navnet på plastokinon stammer faktisk fra plasseringen i kloroplaster av høyere planter.
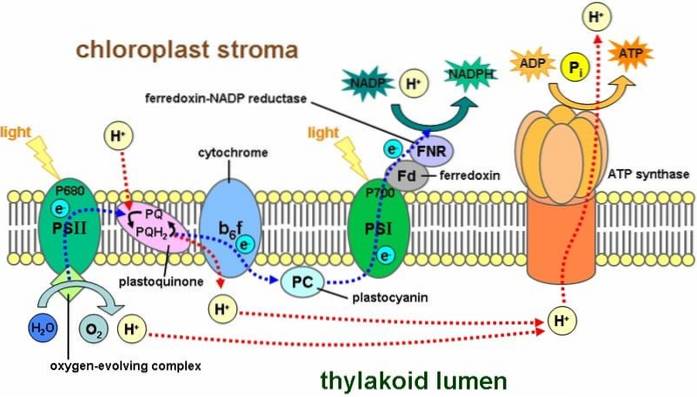
Under fotosyntese fanges solstråling i FS-II-systemet av klorofyll P-680 og oksideres deretter ved frigjøring av et elektron. Denne elektronen stiger til et høyere energinivå, som blir plukket opp av velgerens akseptormolekyl: plastokinon (PQ)..
Plastokinoner er en del av den fotosyntetiske elektrontransportkjeden. De er stedet for integrering av forskjellige signaler og en nøkkelbit i RSp31s respons på lys. Det er rundt 10 PQ per FS-II som reduseres og oksyderes i henhold til den funksjonelle tilstanden til det fotosyntetiske apparatet..
Derfor overføres elektronene gjennom en transportkjede der flere cytokromer er involvert, for senere å nå plastocyanin (PC), som vil gi elektronene til klorofyllmolekyler av FS-I.
Artikkelindeks
Plastokinon (C55H80ELLERto) er et molekyl assosiert med en benzenring (kinon). Spesielt er det en isomer av cykloheksadion, karakterisert ved å være en aromatisk forbindelse differensiert av redokspotensialet.
Kinoner er gruppert basert på struktur og egenskaper. Innenfor denne gruppen er benzokinoner differensiert, generert av oksygenering av hydrokinoner. Isomerer av dette molekylet er orto-benzokinon og for å-benzokinon.
På den annen side ligner plastokinon ubiquinon, fordi de tilhører benzokinonfamilien. I dette tilfellet fungerer begge som elektronakseptorer i transportkjeder under fotosyntese og anaerob respirasjon..
Assosiert med lipidstatus, er den kategorisert i terpenfamilien. Det vil si lipidene som utgjør pigmenter fra planter og dyr som gir celler farge.
Plastokinon består av en aktiv benzen-kinonring assosiert med en sidekjede av et polyisoprenoid. Faktisk er den sekskantede aromatiske ringen knyttet til to oksygenmolekyler gjennom dobbeltbindinger ved C-1- og C-4-karbonene..
Dette elementet har sidekjeden og består av ni isoprener koblet sammen. Følgelig er det en polyterpen eller isoprenoid, det vil si hydrokarbonpolymerer med fem karbonatomer isopren (2-metyl-1,3-butadien).
På samme måte er det et prenylert molekyl som letter feste til cellemembraner, i likhet med lipidankre. I denne forbindelse har en hydrofob gruppe blitt tilsatt til dens alkylkjede (metylgruppe CH3 forgrenet i posisjon R3 og R4).
Under den fotosyntetiske prosessen blir plastokinon kontinuerlig syntetisert på grunn av sin korte livssyklus. Studier i planteceller har bestemt at dette molekylet forblir aktivt mellom 15 og 30 timer.
Faktisk er biosyntese av plastokinon en veldig kompleks prosess som involverer opptil 35 enzymer. Biosyntese har to faser: den første forekommer i benzenringen og den andre i sidekjedene.
I den innledende fasen ble syntesen av kinon-benzenringen og prenylkjeden utført. Ringen oppnådd fra tyrosiner og prenyl-sidekjeder er resultatet av glyseraldehyd-3-fosfat og pyruvat..
Basert på størrelsen på polyisoprenoidkjeden, blir typen plastokinon etablert.
Den neste fasen omfatter kondensasjonsreaksjonen til ringen med sidekjedene.
Homogentistsyre (HGA) er forgjengeren til benzen-kinonringen, som er syntetisert fra tyrosin, en prosess som oppstår takket være katalysen av enzymet tyrosinaminotransferase.
På sin side stammer prenyl-sidekjedene i metylerytritolfosfat (MEP) -veien. Disse kjedene katalyseres av enzymet solanesyldifosfatsyntetase for å danne solanesyldifosfat (SPP).
Metylerytritolfosfat (MEP) utgjør en metabolsk vei for biosyntese av isoprenoider. Etter dannelsen av begge forbindelsene, oppstår kondensering av homogenistisk syre med solanesyldifosfatkjeden, en reaksjon katalysert av enzymet homogentistat solanesyl-transferase (HST).
Til slutt stammer en forbindelse kalt 2-dimetyl-plastokinon, som senere med intervensjonen av enzymet metyltransferase tillater å oppnå som et sluttprodukt: plastokinon.
Plastokinoner griper inn i fotosyntese, en prosess som skjer med inngrep av energi fra sollys, noe som resulterer i energirikt organisk materiale fra transformasjonen av et uorganisk substrat.
Funksjonen til plastokinon er assosiert med lysfasen (PS-II) av den fotosyntetiske prosessen. Plastokinonmolekylene involvert i elektronoverføring kalles QA og QB.
I denne forbindelse er fotosystem II (PS-II) et kompleks som kalles vann-plastokinonoksidreduktase, hvor to grunnleggende prosesser utføres. Vannoksidasjon katalyseres enzymatisk og plastokinonreduksjon oppstår. I denne aktiviteten absorberes fotoner med en bølgelengde på 680 nm..
Molekyler Q A og Q B er forskjellige i måten de overfører elektroner på og hastigheten på overføringen. Også på grunn av typen binding (bindingsside) med fotosystem II. Q A sies å være den faste plastokinon og Q B er den mobile plastokinon.
QA er tross alt bindingssonen for fotosystem II som aksepterer de to elektronene i en tidsvariasjon mellom 200 og 600 oss. I stedet har Q B muligheten til å binde og løsne seg fra fotosystem II, akseptere og overføre elektroner til cytokromet..
På molekylært nivå, når QB er redusert, byttes det mot et annet av settet med gratis plastokinoner i tylakoidmembranen. Mellom Q A og Q B er det et ikke-ionisk Fe-atom (Fe+to) som deltar i elektronisk transport mellom dem.
Kort fortalt samhandler QB med aminosyrerester i reaksjonssenteret. På denne måten får Q A og Q B en stor differensial i redokspotensialene..
Siden QB er mer løst bundet til membranen, kan den dessuten enkelt skilles fra ved å reduseres til QH 2. I denne tilstanden er den i stand til å overføre høyenergielektroner mottatt fra QA til cytokrom bc1-kompleks 8.

Ingen har kommentert denne artikkelen ennå.