De polyploidi Det er en type genetisk mutasjon som består av tilsetning av et komplett sett (komplette sett) av kromosomer til cellekjernen, som utgjør homologe par. Denne typen kromosommutasjon er den vanligste av euploidiene og er preget av det faktum at organismen bærer tre eller flere komplette sett med kromosomer..
En organisme (vanligvis diploid = 2n) regnes som polyploid når den får et eller flere komplette sett med kromosomer. I motsetning til punktmutasjoner, kromosomale inversjoner og dupliseringer, er denne prosessen i stor skala, det vil si at den forekommer på komplette sett med kromosomer.
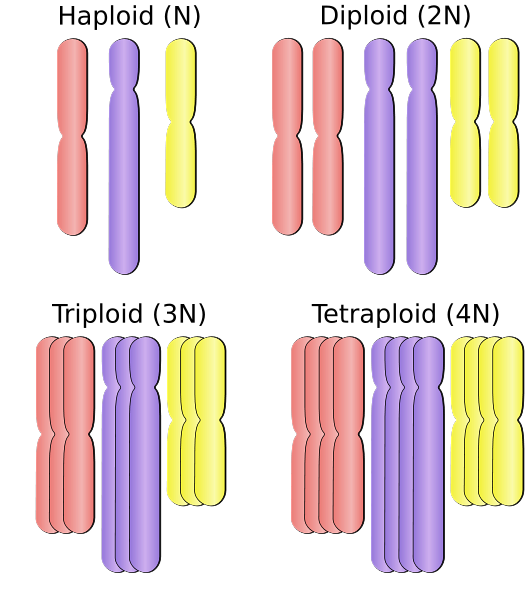
I stedet for å være haploide (n) eller diploide (2n), kan en polyploide organisme være tetraploide (4n), octoploid (8n), eller større. Denne mutasjonsprosessen er ganske vanlig hos planter og er sjelden hos dyr. Denne mekanismen kan øke genetisk variasjon i sittende organismer som ikke er i stand til å bevege seg fra et miljø til et annet..
Polyploidi er av stor betydning evolusjonært i visse biologiske grupper, hvor den utgjør en hyppig mekanisme for generering av nye arter siden kromosombelastning er en arvelig tilstand..
Artikkelindeks
Forstyrrelser i kromosomtall kan forekomme både i naturen og i laboratoriebestemte populasjoner. De kan også induseres med mutagene midler som colchicine. Til tross for meioseens utrolige presisjon, forekommer kromosomavvik og er mer vanlig enn man skulle tro.
Polyploidi oppstår som et resultat av noen endringer som kan oppstå under meiose, enten i den første meiotiske divisjonen eller under profasen, der de homologe kromosomene er organisert i par for å danne tetrader og en ikke-adskillelse av sistnevnte oppstår under anafase I.
Polyploidi er viktig da det er et utgangspunkt for å opprette nye arter. Dette fenomenet er en viktig kilde til genetisk variasjon, siden det gir opphav til hundrevis eller tusenvis av dupliserte lokus som er fri til å skaffe nye funksjoner..
I planter er det spesielt viktig og ganske utbredt. Det anslås at mer enn 50% av blomstrende planter har sitt utspring fra polyploidi.
I de fleste tilfeller skiller polyploider seg fysiologisk fra den opprinnelige arten, og på grunn av dette kan de kolonisere miljøer med nye egenskaper. Mange viktige arter i landbruket (inkludert hvete) er polyploider av hybrid opprinnelse.
Polyploidier kan klassifiseres i henhold til antall sett eller komplette kromosomsett som er tilstede i cellekjernen.
I denne forstand er en organisme som inneholder "tre" sett med kromosomer, "triploid", "tetraploid" hvis den inneholder 4 sett med kromosomer, pentaploid (5 sett), hexaploidae (6 sett), heptaploid (syv sett), blekksprut ( åtte kamper), nonaploidae (ni kamper), decaploid (10 kamper), og så videre.
På den annen side kan polyploidier også klassifiseres etter opprinnelsen til de kromosomale begavelsene. I denne ideen kan en organisme være: autopolyploid eller allopolypoid.
En autopolyploid inneholder flere sett med homologe kromosomer avledet fra samme individ eller fra et individ som tilhører samme art. I dette tilfellet dannes polyploider ved foreningen av ikke-reduserte gameter av genetisk kompatible organismer som er katalogisert som den samme arten..
En allopolypoid er en organisme som inneholder ikke-homologe sett med kromosomer på grunn av hybridisering mellom forskjellige arter. I dette tilfellet forekommer polyploidi etter hybridisering mellom to beslektede arter..
Polyploidi er sjelden eller sjelden hos dyr. Den mest utbredte hypotesen som forklarer den lave frekvensen av polyploide arter i høyere dyr, er at deres komplekse mekanismer for kjønnsbestemmelse avhenger av en veldig delikat balanse i antall kjønnskromosomer og autosomer..
Denne ideen har blitt opprettholdt til tross for akkumulert bevis på dyr som eksisterer som polyploider. Det observeres vanligvis i lavere dyregrupper som ormer og et stort utvalg av flatorm, der individer vanligvis har både mannlige og kvinnelige kjønnsorganer, noe som letter selvgjødsling..
Arter med sistnevnte tilstand kalles selvkompatible hermafroditter. På den annen side kan det også forekomme i andre grupper hvis kvinner kan gi avkom uten befruktning, gjennom en prosess som kalles parthenogenese (som ikke innebærer en normal meiotisk seksuell syklus)
Under parthenogenese produseres avkom i utgangspunktet ved mitotisk deling av forelderceller. Dette inkluderer mange arter av virvelløse dyr som biller, isopoder, møll, reker, forskjellige grupper av arachnids, og noen arter av fisk, amfibier og reptiler..
I motsetning til planter er spesiering gjennom polyploidi en eksepsjonell begivenhet hos dyr.
Gnageren Tympanoctomys barriere det er en tetraploid art som har 102 kromosomer per somatisk celle. Det har også en "gigantisk" effekt på sædceller. Denne allopolyploide arten stammer sannsynligvis fra forekomsten av flere hybridiseringshendelser av andre gnagerarter som Octomys mimax Y Pipanacoctomys aureus.
Polyploidi er uvanlig hos virveldyr og regnes som irrelevant i diversifiseringen av grupper som pattedyr (i motsetning til planter) på grunn av forstyrrelser som oppstår i kjønnsbestemmelsessystemet og i dosekompensasjonsmekanismen..
Det anslås at fem av 1000 mennesker er født med alvorlige genetiske defekter som kan tilskrives kromosomavvik. Enda flere embryoer med kromosomfeil abort, og mange flere kommer aldri til fødselen..
Kromosomale polyploidier regnes som dødelige hos mennesker. Imidlertid er omtrent 50% av disse i somatiske celler som hepatocytter normalt polyploide (tetraploide eller oktaploide)..
De hyppigst oppdagede polyploidiene i vår art er komplette triploider og tetraploider, samt diploide / triploide (2n / 3n) og diploide / tetraploide (2n / 4n) mixoploider..
I sistnevnte eksisterer en populasjon av normale diploide celler (2n) sammen, med en annen som har 3 eller flere haploide multipler av kromosomer, for eksempel: triploide (3n) eller tetraploide (4n).
Triploidier og tetraplodia hos mennesker er ikke levedyktige på lang sikt. Død ved fødselen eller til og med noen få dager etter fødselen har i de fleste tilfeller blitt rapportert, varierende fra mindre enn en måned til maksimalt 26 måneder..
Eksistensen av mer enn ett genom i samme kjerne har spilt en viktig rolle i plantens opprinnelse og evolusjon, og er kanskje den viktigste cytogenetiske endringen i plantespesiering og evolusjon. Planter var inngangsporten til kunnskapen til celler med mer enn to sett med kromosomer per celle.
Fra begynnelsen av kromosomantal ble det observert at et stort utvalg av ville og dyrkede planter (inkludert noen av de viktigste) var polyploid. Nesten halvparten av de kjente artene av angiospermer (blomstrende planter) er polyploid, de fleste bregner (95%) og et bredt utvalg av moser.
Tilstedeværelsen av polyploidi i gymnospermplanter er sjelden og svært variabel i grupper av angiospermer. Generelt har det blitt påpekt at polyploidplanter er svært tilpasningsdyktige, og er i stand til å okkupere habitater som deres diploide forfedre ikke kunne. I tillegg akkumulerer polyploidplanter med flere genomiske kopier større "variabilitet".
Innenfor planter spilte kanskje allopolyploidene (mer vanlig i naturen) en grunnleggende rolle i spesiering og adaptiv stråling fra mange grupper..
I planter kan polyploidi stamme fra flere forskjellige fenomener, kanskje de hyppigste er feil under meioseprosessen som gir opphav til diploide kjønnsceller..
Mer enn 40% av de dyrkede plantene er polyploid, inkludert alfalfa, bomull, poteter, kaffe, jordbær, hvete blant andre, uten et forhold mellom domesticering og polyploidi av plantene..
Siden colchicine ble implementert som et middel for å indusere polyploidi, har det blitt brukt i avlingsplanter av tre grunner:
-Å generere polyploidi hos visse viktige arter, som et forsøk på å oppnå bedre planter, siden det i polyploider vanligvis er en fenotype der det er en betydelig vekst av "gigabyte" på grunn av det faktum at det er et større antall celler. Dette har tillatt bemerkelsesverdige fremskritt innen hagebruk og innen plantegenetisk forbedring..
-For polyploidisering av hybrider og for å gjenvinne fruktbarheten på en slik måte at noen arter blir redesignet eller syntetisert.
-Og til slutt, som en måte å overføre gener mellom arter med forskjellige grader av ploidi eller innenfor samme art..
Innen planter er en naturlig polyploid av stor betydning og spesielt interessant brødhvete, Triticum aestibum (heksaploid). Sammen med rug ble det med vilje bygd en polyploid kalt "Triticale", en allopolyploid med høy produktivitet av hvete og robustheten til rug, som har stort potensial.
Hvete i dyrkede planter har vært utrolig viktig. Det er 14 arter av hvete som har utviklet seg ved allopolypoidi, og de danner tre grupper, en av 14, en annen av 28 og en siste av 42 kromosomer. Den første gruppen inkluderer de eldste artene av slekten T. monococcum Y T. boeoticum.
Den andre gruppen består av 7 arter og kommer tilsynelatende fra hybridiseringen av T. boeoticum med en art av urt av en annen slekt kalt Aegilops. Krysset produserer en kraftig steril hybrid som gjennom kromosom duplisering kan resultere i en fruktbar allotetraploid.
Den tredje gruppen på 42 kromosomer er der hvor brødvete ligger, som sannsynligvis stammer fra hybridisering av en tertraploid art med en annen art av Aegilops etterfulgt av en duplisering av det kromosomale komplementet.

Ingen har kommentert denne artikkelen ennå.