De telofase det er den siste delingsfasen av mitose og meiose. Det er etter anafase og går forut for cytoplasmisk deling eller cytokinese. Det karakteristiske trekket som skiller og definerer det, er dannelsen av nye kjerner.
Når det dupliserte DNA ble komprimert (profase), migrerte de bundne søsterkromatidene til cellens ekvator (metafase). Når de alle var samlet der, stilte de seg opp for å bli mobilisert til polene i delingscellen under anafase..
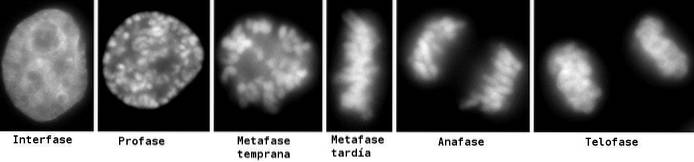
Til slutt, for å dele og gi opphav til to celler, må to kjerner først dannes for å beskytte DNA. Det er nettopp dette som skjer under telofasen av mitose..
Ikke at noe veldig annet skjer, mekanisk sett, under telofasene til meiose I og meiose II. Men materialene som mottas som "kromosomer" er veldig forskjellige..
I telofase I mottar cellen i meiose bare ett sett med dupliserte homologer ved hver pol. Det vil si et enkelt sett av det kromosomale komplementet til arten med hvert kromosom dannet av to søsterkromatider sammen med sentromeren.
Ved telofasen av meiose II migrerer søsterkromatider poleward, og det dannes kjerner med et haplooid antall kromosomer. På slutten av telofasen er kromosomene ikke lenger synlige som komprimerte strukturer.
Artikkelindeks
I denne delen vil vi se på tre definerende aspekter ved telofaser: begynnelsen av dannelsen av nukleoli, dekondensasjonen av kromatin og utseendet til nye kjernekonvolutter..
I åpne mitoser dannes mange små nukleoli, som etter hvert som syklusen utvikler seg, smelter sammen og danner nukleolene som er typiske for arten (som ikke er mange). Med hendelser som ble utløst under metafase, begynner den strukturelle biogenesen av disse organellene i telofase..
Dette er av stor betydning fordi i nukleoli syntetiseres blant annet RNAene som er en del av ribosomene. I ribosomer utføres prosessen med translasjon av messenger-RNA for å produsere proteiner. Og hver celle, spesielt nye, trenger å lage proteiner raskt..
Ved å dele vil derfor hvert nye celleprodukt fra den divisjonen være kompetent for oversettelsesprosessen og den autonome eksistensen.
På den annen side er kromatinet som arves fra anafase, høyt komprimert. Dette må dekondenseres for å organisere det i kjernene i formasjon i åpne mitoser..
Rollen til å kontrollere kromatindekondensering i en delende celle spilles av en proteinkinase kalt Aurora B. Dette enzymet begrenser dekondensasjonsprosessen under anafase, og begrenser den til den siste fasen av deling eller telofase. Faktisk er Aurora B proteinet som styrer overgangen fra anafase til telofase..
Det andre viktige aspektet ved telofasen, og som definerer det, er dannelsen av atomkonvolutten. La oss huske at i åpne celledelinger forsvinner kjernekonvolutten for å tillate en gratis mobilisering av det kondenserte kromatinet. Nå som kromosomene har segregert, må de grupperes i en ny kjerne per cellepol.
For å generere en ny kjerne, må kromatin samhandle med proteinene som vil danne kjernelaminat, eller lamininer. Lamininene vil i sin tur tjene som en bro for interaksjonen med andre proteiner som vil tillate dannelse av kjernelaminat.
Dette vil skille kromatinet inn i eu- og heterokromatin, tillate den indre organisasjonen av kjernen og hjelpe til med konsolidering av den indre kjernemembranen..
Samtidig vil mikrotubuli-strukturer avledet fra stamcellens endoplasmatiske retikulum migrere til kondensasjonssonen til telofasekromatin. De vil dekke det i små flekker, og deretter smelte sammen for å dekke det helt.
Dette er den eksterne kjernemembranen som er kontinuerlig med det endoplasmatiske retikulumet og med den indre kjernemembranen..
Alle de ovennevnte trinnene beskriver telofasen av mitose i grunnleggende grunnlag. Ved hver cellepol vil det dannes en kjerne med det kromosomale komplementet til modercellen..
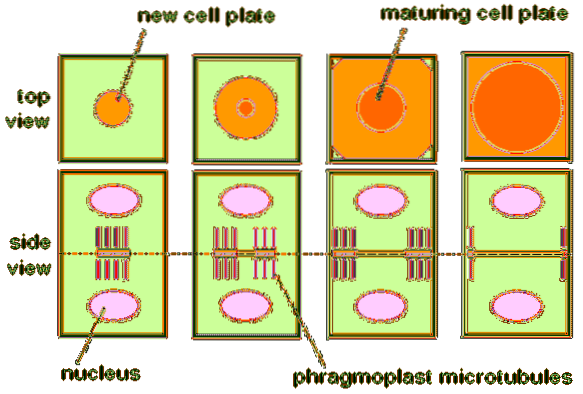
Men i motsetning til mitose hos dyr, dannes det under mitose i planteceller en unik struktur kjent som en fragmoplast. Dette vises mellom de to fremtidige kjernene i overgangen mellom anafase og telofase..
Hovedrollen i plante mitotisk deling er å syntetisere celleplaten. Det vil si at fragmoplasten genererer stedet der de nye cellene i planten vil dele seg når telofasen slutter..
I meiotiske telofaser forekommer det som allerede er beskrevet, men med noen forskjeller. I telofase I dannes "kjerner" med et enkelt sett med homologe (dupliserte) kromosomer. Kjerner dannes i telofase II med et haploid komplement av søsterkromatider.
I mange organismer forekommer ikke kromatinkondensasjon ved telofase I, som overgår nesten umiddelbart til meiose II. I andre tilfeller dekondenserer kromatinet, men det komprimerer seg raskt igjen under profase II..
Atomkonvolutten er vanligvis kortvarig i telofase I, men permanent i II. Aurora B-proteinet kontrollerer segregeringen av homologe kromosomer under telofase I. Imidlertid deltar den ikke i segregeringen av søsterkromatider under telofase II..
I alle tilfeller av kjernedeling blir denne prosessen fulgt av en deling av cytoplasma, en prosess som kalles cytokinese. Cytokinesis observeres både på slutten av telofase i mitose, og på slutten av telofase I og telofase II av meiose.


Ingen har kommentert denne artikkelen ennå.