
Tonoplast er begrepet brukt i biologi for å identifisere de indre membranene i vakuolene i planteceller. Tonoplast har selektiv permeabilitet og låser vann, ioner og oppløste stoffer i vakuoler..
Det er uttømmende studier på den molekylære sammensetningen av tonoplastene, siden transportørproteinene i disse membranene regulerer plantevekst, stress mot saltinnhold og uttørking, og følsomhet for patogener..
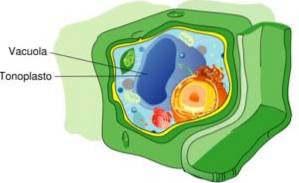
Generelt inneholder vakuolen som tonoplasten består av 57,2% av hele cellevolumet i planter. Imidlertid kan denne prosentandelen variere avhengig av livsstilen, det å være kaktus og ørkenplanter vanligvis de som har mindre eller større vakuoler..
I noen plantearter kan vakuolen avgrenset av tonoplasten oppta opptil 90% av det indre volumet av alle planteceller.
Siden det er involvert i en konstant trafikk av molekyler, ioner og enzymer mellom cytosolen og det indre av vakuolen, er tonoplasten rik på transportørproteiner, kanaler og akvaporiner (porer eller kanaler som vann passerer gjennom).
Mange av de indre vesiklene, som fagosomer eller transportvesikler, smelter sammen med tonoplasten for å avsette innholdet i vakuolen, hvor deres bestanddeler kan nedbrytes og resirkuleres..
Bioteknologer fokuserer sin innsats på teknikkene som er nødvendige for å innlemme tonoplaster i planter av kommersiell interesse som hvete og ris med egenskapene til planter som er motstandsdyktige mot saltoppløsning..
Artikkelindeks
Tonoplasten består for det meste av proteiner og lipider anordnet i form av et lipiddobbeltlag, mer eller mindre lik plasmamembranen til celler. Imidlertid, når det sammenlignes med andre cellemembraner, har den unike proteiner og lipider i sammensetningen..
Den vakuolære membranen (tonoplasten) består av 18% nøytrale lipider og steroler, 31% glykolipider og 51% fosfolipider. Normalt er fettsyrene som er tilstede i lipidene som danner dobbeltlaget fullstendig mettet, det vil si at de ikke har dobbeltbindinger..
Den enorme vakuolen definert av tonoplasten begynner som et sett med flere små vakuoler som syntetiseres i endoplasmatisk retikulum, senere proteiner fra Golgi-apparatet er innlemmet..
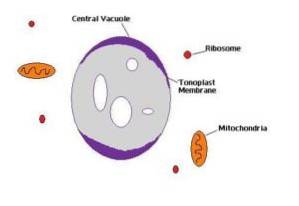
Proteinene fra Golgi-apparatet er kanalene, enzymene, transportøren og strukturelle proteiner og de forankrende glykoproteinene som vil bli plassert i tonoplasten..
Alle de små vakuolene smelter sammen og organiseres sakte og progressivt til de danner tonoplasten som gir opphav til en stor vakuol, hovedsakelig fylt med vann og ioner. Denne prosessen forekommer i alle organismer i riket Plantae, derfor har alle planteceller tonoplast.
Tonoplasten har, i likhet med mitokondrie lipid dobbeltlag, mellom sin struktur to typer primære protonpumper, en ATPase og en pyrofosfatase, som gjør det mulig for det indre av vakuolen å ha en sur pH.
Tonoplastens hovedfunksjon er å fungere som en semipermeabel barriere, avgrense rommet som vakuolen består av, og skille det fra resten av det cytosoliske innholdet..
Denne "semipermeabiliteten" brukes av planteceller for turgor, pH-kontroll, vekst, blant mange andre funksjoner..
Den mest studerte funksjonen til tonoplasten i planter er å regulere celleturgor. Konsentrasjonen av ioner og vann som er funnet inne i vakuolen deltar gjennom trykkpotensialet (Ψp) i vannpotensialet (Ψ) slik at vannmolekylene kommer inn i eller forlater det indre av cellen.
Takket være tilstedeværelsen av tonoplasten genereres trykkpotensialet (Ψp) som utøves av protoplasten (plasmamembranen) på celleveggen i celler. Denne kraften får positive verdier ettersom vakuolen utøver trykk på protoplasten, og dette igjen på celleveggen.
Når vannet forlater vakuolen gjennom tonoplasten og deretter forlater plantecellen, begynner vakuolen å trekke seg sammen og turgoren til cellen går tapt, og oppnår trykkpotensialverdier (Ψp) nær null og til og med negativ..
Denne prosessen er kjent som begynnende plasmolyse og er det som igjen produserer visnen som vi observerer i planter.
Når planten visner, øker det cellulære osmotiske potensialet (Ψp), siden når konsentrasjonen av kaliumioner (K +) inne i cellen er større enn konsentrasjonen av oppløste stoffer utenfor, beveger vannet seg innover..
Disse kaliumionene (K +) finnes hovedsakelig inne i vakuolen og er sammen med cytosolionene ansvarlige for å generere det osmotiske potensialet (Ψp). Tonoplast er gjennomtrengelig for disse kaliumionene takket være en ATPase som den har i sin struktur.
ATPaser i tonoplasten opprettholder en konstant protongradient mellom cytosolen og det indre av vakuolen..
ATPases av rotcellemembranen aktiveres av tilstedeværelsen av kaliumioner (K +), disse introduserer kaliumioner (K +) og utviser protoner (H +). I motsetning til dette aktiveres ATPaser som finnes i tonoplasten i nærvær av klor (Cl-) i cytosolen..
Disse styrer konsentrasjonen av de indre klor- (Cl-) og hydrogen- (H +) -ionene. Begge ATPases fungerer i et slags "spill" for å kontrollere pH i cytosolen til planteceller, enten ved å øke eller senke pH til en pH på 7 eller høyere i cytosolen..
Når det er en veldig høy konsentrasjon av protoner (H +) i cytosolen, introduserer ATPase av cellemembranen kaliumioner (K +); mens ATPase av tonoplast suger klor (Cl-) og hydrogen (H +) ioner fra cytosolen inn i vakuolen.
Tonoplast har flere typer primære protonbomber. I tillegg har den transportkanaler for kalsiumioner (Ca +), hydrogenioner (H +) og andre ioner som er spesifikke for hver planteart..
ATPases pumper protoner (H +) inn i vakuolen, noe som får lumenet til å få en sur pH, med verdier mellom 2 og 5, og en positiv delvis ladning. Disse pumpene hydrolyserer ATP i cytosolen, og gjennom en pore introduserer protoner (H +) i lumenet i vakuolen..
Pyrofosfataser er en annen type tonoplast “pumper” som også introduserer protoner (H +) i vakuolen, men gjør det gjennom hydrolyse av pyrofosfat (PPi). Denne pumpen er unik for planter og avhenger av Mg ++ og K-ioner+.
En annen type ATPaser finnes i tonoplasten som pumper protoner inn i cytosolen og introduserer kalsiumioner (Ca ++) i vakuolen. Kalsium (Ca ++) brukes som et budbringer inne i cellen og lumenet til vakuolen brukes som et reservoar for disse ionene.
Kanskje de mest vanlige proteinene i tonoplasten er kalsiumkanaler, disse tillater utgang av kalsium (Ca +) introdusert av ATPaser i membranen.
For tiden er primærpumper eller transportører av typen ABC (fra engelsk TILTP-Binding Cassette) i stand til å introdusere store organiske ioner i vakuolen (for eksempel glutation, for eksempel).


Ingen har kommentert denne artikkelen ennå.