De tubulin er et kuleformet dimerprotein som består av to polypeptider: alfa og beta tubulin. De er ordnet i form av et rør for å gi opphav til mikrotubuli, som sammen med aktinmikrofilamenter og mellomfilamenter utgjør cytoskelettet.
Mikrotubuli finnes i forskjellige viktige biologiske strukturer, for eksempel flagell av sædceller, utvidelser av ciliated organismer, cilia i luftrøret og eggleder, blant andre..
I tillegg fungerer strukturene som tubulin danner som transportveier - analoge med togsporene - av materialer og organeller i cellen. Forskyvning av stoffer og strukturer er mulig takket være motoriske proteiner assosiert med mikrotubuli, kalt kinesin og dynein..
Artikkelindeks
Tubulin-underenheter er 55.000 dalton heterodimerer og er byggesteinene til mikrotubuli. Tubulin finnes i alle eukaryote organismer og har vært svært konservert gjennom evolusjonen.
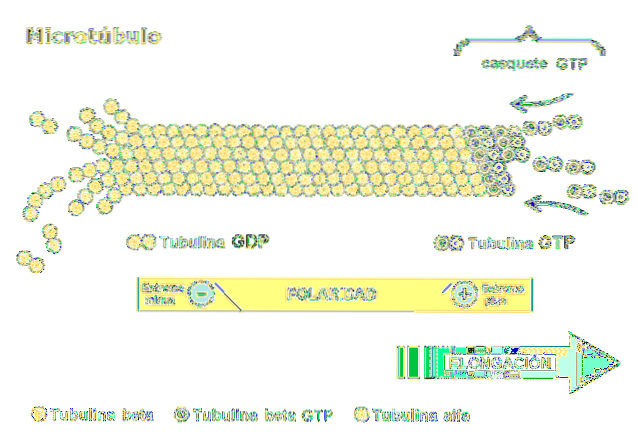
Dimeren består av to polypeptider kalt alfa og beta tubulin. Disse polymeriserer for å danne mikrotubuli, som består av tretten protofilamenter anordnet parallelt i form av et hulrør..
En av de mest relevante egenskapene til mikrotubuli er strukturens polaritet. Med andre ord er de to endene av mikrotubuli ikke de samme: den ene enden kalles den raskt voksende eller "mer" enden, og den andre er den langsomt voksende eller "mindre" enden..
Polaritet er viktig ettersom den bestemmer bevegelsesretningen langs mikrorøret. Tubulin dimer er i stand til å polymerisere og depolarisere i raske monteringssykluser. Dette fenomenet forekommer også i aktinfilamenter.
Det er en tredje type underenhet: det er gammatubulin. Dette er ikke en del av mikrotubuli og ligger i sentrosomene; imidlertid deltar den i kjerneformering og dannelse av mikrotubuli.
Alfa- og beta-underenhetene assosierer sterkt for å danne en kompleks heterodimer. Faktisk er samspillet mellom komplekset så intenst at det ikke dissosierer under normale forhold..
Disse proteinene består av 550 aminosyrer, for det meste sure. Selv om alfa- og beta-tubuliner er ganske like, blir de kodet av forskjellige gener..
I alfatubulin kan aminosyrerester med en acetylgruppe bli funnet, noe som gir den forskjellige egenskaper i celleflageller..
Hver underenhet av tubulin er assosiert med to molekyler: i alfa-tubulin binder GTP irreversibelt og hydrolyse av forbindelsen forekommer ikke, mens det andre bindingsstedet i beta-tubulin binder GTP reversibelt og hydrolyserer det..
GTP-hydrolyse resulterer i et fenomen som kalles "dynamisk ustabilitet" der mikrotubuli gjennomgår vekst- og nedgangssykluser, avhengig av hastigheten på tilsetning av tubulin og hastigheten på GTP-hydrolyse..
Dette fenomenet resulterer i en høy omsetningshastighet for mikrotubuli, hvor strukturens halveringstid bare er noen få minutter..
Alfa- og beta-underenhetene til tubulin polymeriserer for å gi opphav til mikrotubuli, som er en del av cytoskelettet.
I tillegg til mikrotubuli består cytoskjelettet av to ekstra strukturelle elementer: aktinmikrofilamenter på omtrent 7 nm og mellomfilamenter med en diameter på 10 til 15 nm..
Cytoskjelettet er rammen til cellen, den støtter og opprettholder celleformen. Imidlertid er membran- og subcellulære rom ikke statiske og er i konstante bevegelser for å kunne utføre fenomenene endocytose, fagocytose og utskillelse av materialer..
Strukturen til cytoskjelettet lar cellen tilpasse seg selv for å oppfylle alle de nevnte funksjonene..
Det er det ideelle mediet for celleorganeller, plasmamembranen og andre cellekomponenter å utføre sine normale funksjoner, i tillegg til å delta i celledeling.
De bidrar også til cellulære bevegelsesfenomener som amoeba-bevegelse, og spesialiserte strukturer for bevegelse som cilia og flagella. Endelig er det ansvarlig for bevegelsen av musklene.
Takket være dynamisk ustabilitet kan mikrotubuli omorganiseres fullstendig under celledelingsprosesser. Mikrotubuli-arrangementet under grensesnittet er i stand til å demonteres, og tubulinunderenhetene er gratis.
Tubulin kan montere seg selv og gi opphav til den mitotiske spindelen, som er involvert i separasjonen av kromosomer.
Det er visse medisiner, som colchicine, taxol og vinblastine som forstyrrer prosessene med celledeling. Den virker direkte på tubulinmolekyler, og påvirker fenomenet montering og dissosiasjon av mikrotubuli.
I dyreceller strekker mikrotubuli seg til sentrosomet, en struktur nær kjernen som består av et par sentrioler (hver orientert vinkelrett) og er omgitt av en amorf substans, kalt pericentriolar matrix..
Sentriolene er sylindriske legemer dannet av ni tripletter av mikrotubuli, i en organisasjon som ligner cilia og celleflagell..
I prosessen med celledeling strekker mikrotubuli seg fra sentrosomene, og danner dermed den mitotiske spindelen, som er ansvarlig for riktig fordeling av kromosomene til de nye dattercellene..
Det ser ut til at sentriolene ikke er essensielle for montering av mikrotubuli i celler, siden de ikke er tilstede i planteceller eller i noen eukaryote celler, som i ovulene til visse gnagere..
I den pericentriolar matrisen skjer initieringen for montering av mikrotubuli, hvor kjerneformering skjer ved hjelp av gammatubulin.
De tre typene tubulin (alfa, beta og gamma) er kodet av forskjellige gener og er homologe med et gen som finnes i prokaryoter som koder for et 40.000-daltonprotein, kalt FtsZ. Bakterieprotein er funksjonelt og strukturelt lik tubulin.
Det er sannsynlig at proteinet hadde en forfedres funksjon i bakterier og ble modifisert under evolusjonære prosesser, og konkluderte i et protein med funksjonene det utfører i eukaryoter..



Ingen har kommentert denne artikkelen ennå.