
Forestillingen om ekstrapyramidal tilnærming eller det ekstrapyramidale systemet (SEP) oppsto som et resultat av anatomiske og fysiologiske studier med sikte på å forstå hvordan sentralnervesystemet kontrollerte aktiviteten til skjelettmuskulaturen, med sikte på at kroppen antok riktig kroppsholdning og produserte den frivillige bevegelser.
I denne prosessen ble det oppdaget at kontrollen av muskelaktivitet krevde kontroll av motorneuronene i det fremre hornet i ryggmargen, den eneste forbindelsen mellom sentralnervesystemet og skjelettmuskelfibre, og at denne kontrollen ble utøvd av nerveprojeksjoner fra hjernesentre. overlegen.
Blant disse projeksjonene er en viktig vei dannet av noen aksoner som har sitt utspring i motorområdene i hjernebarken og kommer direkte ned, det vil si uten skalaer, til ryggmargen, og blir sammen når de passerer gjennom medulla oblongata, i noen fremtredende som på grunn av sin form fikk navnet "pyramider".
Denne kanalen ble kalt "pyramidekanalen" eller "kortikospinalkanalen", og den var involvert i kontrollen av de fine og dyktige bevegelsene som ble utført av de distale delene av lemmer, mens eksistensen av strukturer med motorisk funksjon ble anerkjent, men ikke inkludert. på denne måten (ekstra).
Begrepet "ekstrapyramidalt motorisk system", som allerede er foreldet fra et fysiologisk synspunkt, brukes fortsatt i klinisk sjargong for å referere til de strukturene i hjernen og hjernestammen som samarbeider om motorisk kontroll, men som ikke er en del av det pyramidale systemet eller direkte kortikospinal.
Artikkelindeks
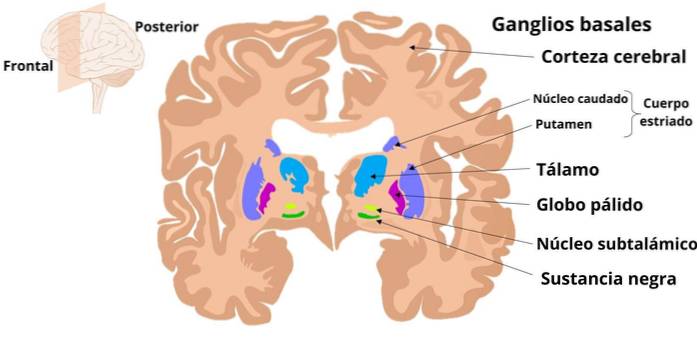
Den ekstrapyramidale banen kan beskrives som organisert i to grupper av komponenter: den ene består av et sett med kjerner i hjernestammen og dets projeksjoner mot ryggmargen, og den andre består av de subkortikale kjernene kjent som kjerner eller basalganglier..
I hjernestammen er det grupper av nevroner hvis aksoner projiserer mot den grå substansen i ryggmargen, og som er beskrevet som organisert i to systemer: det ene medialt og det andre lateralt..
Det mediale systemet er dannet av vestibulospinal, retikulospinal og tektospinal kanal som stiger ned gjennom ledningens ventrale ledninger og utøver kontroll over aksial- eller koffertmusklene, i tillegg til de proksimale musklene i ekstremiteter som er involvert i kroppsholdning..
Den viktigste komponenten i lateralsystemet er rubrospinal-kanalen, hvis aksoner rager ut fra den røde kjernen i midthjernen, kommer ned gjennom ledningens laterale ledning og ender med å påvirke motorneuronene som styrer ekstreme ekstremiteter..
Fra det foregående kan det utledes at det mediale systemet samarbeider om de grunnleggende posturale justeringene, som er nødvendige for frivillig motorisk aktivitet, mens lateralsystemet, sammen med den direkte kortikospinalveien, handler om bevegelsene til ekstremitetene rettet mot et formål som nå og manipulere gjenstander.
Basalganglier er subkortikale nevronale strukturer som er involvert i prosessering av motorinformasjon som planlegging og programmering av komplekse dyktige bevegelser, og hvis endringer gir kliniske manifestasjoner som er gruppert i syndromer kjent som "ekstrapyramidal".
Ganglia inkluderer striatum, som består av putamen og caudatkjernen; den bleke kloden, som har en ytre del (GPe) og en indre del (GPi); substantia nigra, organisert i en kompakt del (SNc) og en retikulert del (SNr), og den subtalamiske eller Lewis-kjernen.
Disse strukturene fungerer ved å motta informasjon hovedsakelig fra forskjellige regioner i hjernebarken; informasjon som aktiverer interne kretsløp som påvirker en utgangs neuronal aktivitet som returnerer via den motoriske delen av thalamus til hjernebarken.
Informasjon om ganglier kommer inn gjennom striatum (caudate og putamen). Derfra starter baner som forbinder seg med utgangskjernene som er GPi og SNr, hvis aksoner går til de ventroanterior og ventrolaterale kjernene i thalamus, som igjen projiserer til cortex.
De forskjellige trinnene i kretsen dekkes av nevroner som tilhører et bestemt nevrokjemisk system og som kan ha en hemmende eller eksitatorisk effekt. De kortikostripede forbindelsene, de thalamisk-kortikale og de subthalamiske fibrene frigjør glutamat og er eksitatoriske.
Nevroner hvis aksoner går ut av striatum bruker gammaaminosmørsyre (GABA) som den viktigste nevrotransmitteren og er hemmende. Det er to subpopulasjoner: den ene syntetiserer stoffet P som samsender [GABA (+ Subst. P)] og det andre enkefalin [GABA (+ Encef.)].
GABA (+ Sust. P) nevroner har D1 dopaminreseptorer og blir begeistret av dopamin (DA); de etablerer også en direkte hemmende forbindelse med basalganglia-utløpene (GPi og SNr) som også er GABAergiske men "+ dynorfin" og hemmer glutamatergiske celler i thalamisk-kortikal projeksjon.
GABA (+ Encef.) Nevroner har dopamin D2-reseptorer og hemmes av dopamin. De etablerer indirekte eksitatorisk forbindelse med utgangene (GPi og SNr), siden de projiserer til GPe som hemmer dens GABAergiske nevroner, som hemmer de glutamaterge nevronene i den subthalamiske kjernen, hvis funksjon er å aktivere utgangene (GPi og SNr).
Den kompakte delen av substantia nigra (SNc) har dopaminerge nevroner (DA) som forbinder med striatum-forbindelser, som allerede nevnt, eksitatorisk D1 på GABA-celler (+ Sub. P) og hemmende D2 på GABA-celler (+ Encef.).
Deretter, og i samsvar med det ovennevnte, ender en aktivering av den direkte banen med å hemme utgangene fra basalganglier og frigjøre aktiviteten i thalamisk-kortikale forbindelser, mens aktivering av den indirekte banen aktiverer utgangene og reduserer thalamisk aktivitet. -kortikal.
Selv om interaksjonene og den nøyaktige fellesfunksjonen til de direkte og indirekte banene som er vurdert, ikke er avklart, hjelper den beskrevne anatomiske og nevrokjemiske organisasjonen oss til å forstå, i det minste delvis, noen patologiske tilstander som skyldes dysfunksjon i basale ganglier.
Selv om de patologiske prosessene som bosetter seg i basale ganglier, er av ulik karakter og påvirker ikke bare visse motoriske funksjoner, men også kognitive, assosiative og emosjonelle funksjoner, har motoriske endringer i kliniske bilder en fremtredende plass, og det meste av forskningen har fokusert på dem..
Bevegelsesforstyrrelsene som er typiske for dysfunksjon i basalganglier kan klassifiseres i en av tre grupper, nemlig:
- Hyperkinesier, som Huntingtons sykdom eller chorea og hemibalisme.
- Hypokinesier, som Parkinsons sykdom.
- Dystonier, som atetose.
Generelt kan det sies at hyperkinetiske forstyrrelser, preget av overdreven motorisk aktivitet, har en reduksjon i hemming som utgangene (GPi og SNr) utøver på thalamisk-kortikale projeksjoner, som blir mer aktive.
Hypokinetiske forstyrrelser ledsages derimot av en økning i denne hemmingen, med en reduksjon i thalamisk-kortikal aktivitet.
Det er en hyperkinetisk lidelse preget av ufrivillig og krampaktig tilfeldig risting i ekstremiteter og orofacial region, koreiform eller "dans" bevegelser som gradvis øker og inhabiliserer pasienten, taleforstyrrelse og progressiv utvikling av demens.
Sykdommen ledsages tidlig av en degenerasjon av GABA (+ Encef.) Striatale nevroner i den indirekte banen.
Siden disse nevronene ikke lenger hemmer GPe GABAergiske nevroner, inhiberer de overdreven den subthalamiske kjernen, som slutter å begeistre de inhiberende utgangene (GPi og SNr), og de thalamisk-kortikale projeksjonene er inhibert..
Den består av de voldsomme sammentrekningene av de proksimale musklene i lemmene, som projiseres med kraft i bevegelser med stor amplitude. Skaden i dette tilfellet er degenerasjonen av den subthalamiske kjernen, noe som resulterer i noe som ligner på det som er beskrevet for chorea, men ikke ved hyperinhibering, men ved ødeleggelse av den subthalamiske kjernen.
Det er preget av vanskeligheter og forsinkelse i initiering av bevegelser (akinesia), bevegelseshemmende (hypokinesi), uttrykksløst ansikts- eller ansiktsuttrykk i en maske, gangendring med redusert tilhørende bevegelse av lemmer under bevegelse og skjelving ufrivillig lem i ro.
Skaden består i dette tilfellet av degenerasjonen av det nigrostriatale systemet, som er de dopaminerge projeksjonene som starter fra den kompakte regionen av substantia nigra (SNc) og forbinder med de striatale nevronene som gir opphav til de direkte og indirekte veiene..
Undertrykkelsen av eksitasjonen som de dopaminerge fibrene utøver på GABA (+ Sust. P) -cellene i den direkte banen, fjerner inhiberingen som disse utøver på GABAergiske utløp (GPi og SNr) mot thalamus, som nå er mer hemmet. intensitet. Det er da en disinhibisjon av utgangene.
På den annen side frigjør undertrykkelse av den inhiberende aktiviteten som dopamin utøver på GABA-cellene (+ Encef.) Av den indirekte banen og øker inhiberingen som de utøver på GABA-cellene til GPe, som inhiberer nevronene i nucleus subthalamic, som deretter hyperaktiverer utgangene.
Som man kan se, er det endelige resultatet av effekten av dopaminerg degenerasjon på de to indre banene, direkte og indirekte, det samme, enten det er desinhibering eller stimulering av GABAergic-utgangene (GPi og SNr) som hemmer kjernene thalamus og reduserer deres produksjon i cortex, noe som forklarer hypokinesen


Ingen har kommentert denne artikkelen ennå.